آلية التوحد
تنتج أعراض التوحد (بالإنجليزية: Autism)عن التغيرات المرتبطة بالنضج في مختلف أنظمة الدماغ. آلية حدوث التوحد ليست مفهومة جَيِّدًا. يمكن تقسيم آليتها إلى مجالين: الفيزيولوجيا المرضية لهياكل الدماغ والعمليات المرتبطة بالتوحد، وعلم النفس العصبي الذي يربط بين هياكل الدماغ والسلوكيات.[1] يبدو أن السلوكيات لديها العديد من الفيزيولوجيا المرضية.[2][3]
وهناك دليل على أن اضطرابات المحور الدماغي المعوي قد تنطوي على ذلك.[4][5][6] مراجعة عام 2015 عرضت اقتراح أن خلل التنظيم المناعي، والتهاب الجهاز الهضمي، وخلل في الجهاز العصبي اللاإرادي، وتغييرات النبيت جرثومي معوي، والإيض الغذائي قد يسبب التهاب الأعصاب في الدماغ والخلل الوظيفي.[4] مراجعة عام 2016 تَوَصَّلَت إلى أن تشوهات الجهاز العصبي المعوي قد تلعب دورًا في الاضطرابات العصبية مثل التوحد. الوصلات العصبية والجهاز المناعي هو مسار قد يسمح للأمراض التي تنشأ في الأمعاء بالانتشار إلى الدماغ.[5]
الكثير من الأدلة تشير إلى اختلال المشبك العصبي كسبب للتوحد.[7] قد تؤدي بعض الطفرات النادرة إلى التوحد عن طريق تعطيل بعض المسارات المشبكية، مثل تلك المرتبطة بالتصاق الخلايا.[8] تشير دراسات استبدال الجينات في الفئران إلى أن أعراض التوحد ترتبط ارتباطًا وثيقًا بخطوات التطور اللاحقة التي تعتمد على النشاط في المشابك العصبية والتغيرات المعتمدة على النشاط.[9] يبدو أن جميع الماسخات المعروفة (العوامل التي تسبب تشوهات خلقية) المتعلقة بخطر التوحد تحدث خلال الأسابيع الثمانية الأولى من الحمل، وعلى الرغم من أن هذا لا يستبعد إمكانية بدء التوحد أو الإصابة به لاحقًا، فهناك دليل قوي على أن التوحد ينشأ في وقت مبكر جدا في التنمية.[10]
الفيزيولوجيا المرضية
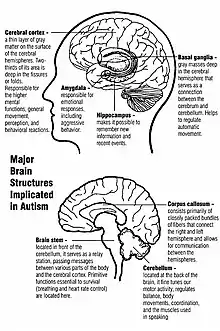
على عكس العديد من اضطرابات الدماغ الأخرى، مثل: مرض باركنسون، التوحد لا يمتلك آلية موحدة واضحة سواء على المستوى الجزيئي أو الخلوي. ومن غير المعروف ما إذا كان التوحد هو عدد قليل من الاضطرابات الناجمة عن الطفرات التي تتلاقى على بضعة مسارات جزيئية مشتركة، أو أنه (مثل التخلف العقلي) مجموعة كبيرة من الاضطرابات ذات آليات متنوعة.[11] يبدو أن التوحد ناتج عن عوامل النمو التي تؤثر على العديد من أنظمة الدماغ الوظيفية أو جميعها،[12] الذي يؤدي إلى تأخير تطور الدماغ أكثر من المنتج النهائي.[13] تشير الدراسات التشريحية العصبية والارتباطات بالماسخات إلى أن آلية التوحد تشمل تغيير نمو الدماغ بعد الحمل مباشرة.[10] ويبدو أن هذا الشذوذ يبدأ سلسلة من الأحداث المرضية في الدماغ التي تتأثر بشكل كبير بالعوامل البيئية.[14] بعد الولادة مباشرة، تميل أدمغة الأطفال المصابين بالتوحد إلى النمو بشكل أسرع من المعتاد، يليها نمو طبيعي أو بطيء نسبيًا في مرحلة الطفولة. ومن غير المعروف ما إذا كان فرط النمو المبكر يحدث في جميع الأطفال المصابين بالتوحد. يبدو أنه أكثر بروزًا في مناطق الدماغ الكامنة وراء تطوير التخصص المعرفي العالي.[15] تتضمن فرضيات الأسس الخلوية والجزيئية للنمو المفرط المرضي المبكر ما يلي:
- وجود فائض من الخلايا العصبية التي تسبب اتصال موضوعي مفرط في مناطق الدماغ الرئيسة.[16]
- هجرة عصبية مضطربة أثناء الحمل المبكر.[17][18]
- عدم توازن في الشبكات المثيرة-المثبطة.[18]
- تشكيل غير طبيعي في المشبك العصبي والشوكات التَغَصُّنِيَّة،[18]على سبيل المثال، عن طريق تعديل نظام التصاق الخلايا نيوركسين-نيوروليجين،[19] أو تنظيم سيء للتخليق الحيوي للبروتين.[20][21] وقد يساهم أيضا تعطل نمو المشبك العصبي في الصرع، مما قد يفسر سبب ارتباط الشرطين.[22]
يُعتقد أن الجهاز المناعي يلعب دورًا مهمًا في التوحد. وجد الباحثون أن الأطفال المصابين بالتوحد لديهم التهاب في كل من الجهاز المناعي المحيطي والمركزي كما يتضح من زيادة مستويات السيتوكينات المؤيدة للالتهابات والتنشيط الكبير للخلايا الدبقية الصغيرة.[23][24][25] كما ارتبطت الدلالة الحيوية للوظيفة المناعية غير الطبيعية بزيادة العجز في السلوكيات التي تتميز بها السمات الأساسية للتوحد مثل: العجز في التفاعلات الاجتماعية والاتصال.[24] تبدأ التفاعلات بين الجهاز المناعي والجهاز العصبي في وقت مبكر خلال المرحلة الجنينية من الحياة، ويعتمد النمو العصبي الناجح على استجابة مناعية متوازنة. يُعتقد أن تنشيط الجهاز المناعي للأم الحامل مثل من المواد السامة البيئية أو العدوى يمكن أن تساهم في إحداث التوحد من خلال التسبب في تعطيل نمو الدماغ.[26][27][28] وتدعم ذلك الدراسات الحديثة التي وجدت أن العدوى أثناء الحمل ترتبط بزيادة خطر الإصابة بالتوحد.[29][30]
إن علاقة المواد الكيميائية العصبية بالتوحد ليست مفهومة جيدًا؛ حُقِّقَ في العديد منها، مع وجود معظم الأدلة على دور السيروتونين والاختلافات الجينية في نقله.[7] أدى الدور الذي تلعبه المجموعة الأولى من مستقبلات الغلوتامات المتحولة (mGluR) في التسبب في متلازمة الكروموسوم إكس الهش، وهي أكثر الجينات شيوعًا في كونها سبب التوحد، إلى الاهتمام بالتداعيات المحتملة في أبحاث مرض التوحد المستقبلة في هذا المسار.[31] وتشير بعض البيانات إلى أن فرط نمو الخلايا العصبية يحتمل أن يكون مرتبطًا بزيادة في عدد من هرمونات النمو[32] أو بضعف تنظيم مستقبلات عوامل النمو. ترتبط بعض الأخطاء الأيضية الخلقية بالتوحد، ولكن من المحتمل أن تكون أقل من 5% من الحالات.[33]
تفترض نظرية نظام الخلايا العصبية المرآتية (MNS) للتوحد أن التشويه في تطور الخلايا العصبية ألمرآتية يتداخل مع التقليد ويؤدي إلى السمات الأساسية للتوحد المتمثلة في الضعف الاجتماعي وصعوبة التواصل. تعمل الخلايا العصبية ألمرآتية عندما يؤدي حيوان عملًا ما أو يلاحظ حيوانا آخرًا يؤدي العمل نفسه. ويمكن أن تساهم هذه الخلايا في فهم الفرد للآخرين وذلك من خلال تمكينه من نمذجة سلوكهم عن طريق محاكة تجسد أفعالهم، ونواياهم ومشاعرهم.[34] وقد اخْتَبَرَت العديد من الدراسات هذه الفرضية من خلال إظهار التشوهات الهيكلية في مناطق الخلايا العصبية المرآتية للأفراد المصابين باضطراب طيف التوحد (ASD)، والتأخر في تنشيط الدائرة الأساسية للتقليد في الأفراد المصابين بمتلازمة أسبرجر، والارتباط بين انخفاض نشاط الخلايا العصبية المرآتية وشدة المتلازمة لدى الأطفال الذين يعانون من اضطراب طيف التوحد (ASD).[35] ومع ذلك، فإن الأفراد المصابين بالتوحد لديهم أيضًا نشاط غير طبيعي للدماغ في العديد من الدوائر خارج MNS[36] ولا تشرح نظرية MNS الأداء الطبيعي للأطفال المصابين بالتوحد في مهام التقليد التي تنطوي على هدف أو كائن.[37]
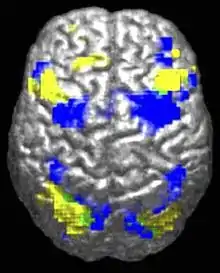
تختلف أنماط التنشيط المنخفض أو الشاذ في الدماغ المرتبطة باضطراب طيف التوحد اعتمادًا على ما إذا كان الدماغ يقوم بمهام اجتماعية أو غير اجتماعية.[38] وفي مجال التوحد هناك أدلة على انخفاض التواصل الوظيفي للشبكة الافتراضية (شبكة دماغية واسعة النطاق تشارك في المعالجة الاجتماعية والعاطفية)، مع الربط السليم للشبكة الإيجابية للمهام (المستخدمة في الاهتمام المستمر والتفكير الموجه نحو الأهداف). وفي حالة المصابين بالتوحد، لا ترتبط الشبكتان سلبًا في الوقت الناسب، مما يشير إلى خلل في تبديل الوظائف بين الشبكتين، وربما يعكس ذلك اضطراب الفكر المرجعي الذاتي.[39]
نظرية نقص الارتباط بالتوحد تفترض أن التوحد يتميز بنقص في الوصلات العصبية العالية المستوى والتزامن، إلى جانب زيادة في العمليات المنخفضة المستوى.[40] عُثِرَ على أدلة لهذه النظرية في دراسات التصوير العصبي الوظيفي للأفراد المصابين بالتوحد[41] ومن خلال دراسة موجات الدماغ التي أشارت أن البالغين المصابين باضطراب طيف التوحد لديهم اتصال موضوعي مفرط في القشرة المخية ووضعف الروابط الوظيفية بين الفص الجبهي وبقية القشرة.[42] تشير أدلة أخرى إلى أن قلة الاتصال يكون بشكل أساسي داخل كل نصف كرة من القشرة وأن التوحد هو اضطراب في القشرة الترابطية.[43]
من الدراسات المستندة إلى الإمكانات المتعلقة بالحدث، والتغييرات العابرة في النشاط الكهربائي للدماغ استجابةً للمحفزات، هناك أدلة كبيرة على وجود اختلافات في الأفراد المصابين بالتوحد فيما يتعلق بالانتباه والتوجه إلى المحفزات السمعية والبصرية واكتشاف الحداثة واللغة ومعالجة الوجه، وتخزين المعلومات؛ وقد وجدت عدة دراسات تفضيلا للمحفزات غير الاجتماعية.[44] على سبيل المثال، وجدت دراسات تخطيط الدماغ المغناطيسي أدلة لدى الأطفال المصابين بالتوحد على تأخر الاستجابة في معالجة الدماغ للإشارات السمعية.[45]
وفي المجال الوراثي، تم العثور على علاقات بين التوحد والفصام استناداً إلى تكرار وحذف الكروموسومات؛ أظهرت الأبحاث أن مرض انفصام الشخصية والتوحد أكثر شيوعًا بشكل ملحوظ مع متلازمة حذف 1q21. إن الأبحاث المتعلقة بالعلاقات بين التوحد والفصام بالنسبة للكروموسوم 15 (15q13.3)، والكروموسوم 16 (16p13.1)، والكروموسوم 17 (17p12) غير حاسمة.[46]
وجدت دراسات الربط الوظيفي كلا من ضعف وفرط الاتصال في أدمغة الأشخاص المصابين بالتوحد. يبدو أن ضعف الاتصال هو المسيطر، خاصةً بالنسبة للتوصيل الوظيفي بين نصفي الكرة والقشرة.[47]
علم النفس العصبي
تم اقتراح فئتين رئيسيتين من النظريات المعرفية حول الروابط بين أدمغة التوحد والسلوك. الفئة الأولى تركز على العجز في الإدراك الاجتماعي.تفترض نظرية التعاطف والتنظيم (ES) لسيمون بارون كوين أن الأفراد المصابين بالتوحد يمكن أن ينظموا—أي أنه يمكنهم تطوير قواعد تشغيل داخلية لمعالجة الأحداث داخل الدماغ—لكنهم أقل فعالية في التعاطف من خلال التعامل مع الأحداث التي تولدها العوامل الأخرى. امتداد، نظرية الدماغ الذكوري المتطرفة، تفترض أن التوحد هو حالة متطرفة من دماغ الذكر، يعرّف نَفْسِيًّا على أنهم الأفراد الذين يكون التنظيم فيهم أفضل من التعاطف.[48] وترتبط هذه النظريات إلى حد ما بنظرية بارون كوين السابقة عن العقل، والتي تفترض أن السلوك التوحدي ينشأ عن عدم القدرة على وصف الحالات الذهنية للنفس وللآخرين. إن نظرية العقل تدعمها الاستجابات غير النمطية للأطفال المصابين بالتوحد لاختبار سالي - آن للتفكير حول دوافع الآخرين،[48] ومن خلال نظام مرآة الخلايا العصبية للتوحد، التي تم وصفها في الخرائط الفيزيولوجية بشكل جيد ومناسب للفرضية.[35] ومع ذلك، لم تجد معظم الدراسات دليلًا على ضعف قدرة الأفراد المصابين بالتوحد على فهم نوايا الآخرين أو أهدافهم الأساسية؛ وبدلًا من ذلك، تشير البيانات إلى أن الإعاقات توجد في فهم العواطف الاجتماعية الأكثر تعقيدًا أو في النظر إلى آراء الآخرين.[49]
وتركز الفئة الثانية على المعالجة الاجتماعية أو المعالجة العامة: الوظائف التنفيذية مثل عمل الذاكرة، التخطيط، والتثبيط. وصرح كنوورثي في استعراضه بأن«ادعاء اختلال الوظائف التنفيذية باعتباره عاملًا مسببًا لمرض التوحد، هو أمر مثير للجدل»، ولكن، «من الواضح أن اختلال الوظائف التنفيذية له دور في العجز الاجتماعي والمعرفي الملحوظ في حالات المصابين بالتوحد».[50] وتشير الاختبارات الخاصة بالوظائف التنفيذية الأساسية مثل مهام حركة العين، إلى وجود تحسن يبدأ في وقت متأخر من مرحلة الطفولة إلى مرحلة المراهقة، ولكن الأداء لا يصل أبدًا إلى المستويات التقليدية التي يصل إليها البالغين.[51] وتتوقع إحدى نقاط النظرية الهامة إلى وجود سلوك نمطي واهتمامات ضيقة؛[52] وهناك نقطتان ضعف لهذه النظرية هما أن الوظيفة التنفيذية يصعب قياسها،[50] وأن العجز في الوظيفة التنفيذية لم يتواجد في حالة الأطفال الصغار المصابين بالتوحد.[53]
نظرية التماسك المركزي الضعيفة (WCC)، تفترض أن القدرة المحدودة على رؤية الصورة الكبيرة هي السبب في الاضطراب المركزي في التوحد. تتمثل إحدى نقاط القوة في هذه النظرية في توقع المواهب الخاصة وذروة الأداء لدى المصابين بالتوحد.[54] وتركز نظرية ذات صلة —تسمى: الأداء الإدراكي المعزز— بشكل أكبر على التفوق في العمليات الموجهة محلياً والعمليات الإدراكية عند الأشخاص الذين يعانون من التوحد.[55] مع ذلك، هناك نوع آخر، وهو monotropism، يفترض أن التوحد ينبع من أسلوب معرفي مختلف، يميل إلى تركيز الانتباه (أو موارد المعالجة) بشكل مكثف، وإلى استبعاد المحفزات الأخرى.[56] هذه النظريات ترسم خريطة جيدة من نظرية ضعف الاتصال للتوحد.
لا تعتبر أي من الفئتين مرضية في حد ذاتها؛ تعالج نظريات الإدراك المعرفي سلوكيات المصابين بالتوحد الجامدة والمتكررة بشكل سيء، بينما تواجه النظريات غير الاجتماعية صعوبة في شرح الضعف الاجتماعي وصعوبات في التواصل.[57] وقد تكون النظرية المشتركة القائمة على حالات العجز المتعددة أكثر فائدة.[58]
المراجع
- "Neurobiological correlates of autism: a review of recent research"، Child Neuropsychol، 12 (1): 57–79، 2006، doi:10.1080/09297040500253546، PMID 16484102، S2CID 46119993.
- "The role of the neurobiologist in redefining the diagnosis of autism"، Brain Pathol، 17 (4): 408–11، 2007، doi:10.1111/j.1750-3639.2007.00103.x، PMID 17919126، S2CID 24860348.
- "Diagnosis of autism"، BMJ، 327 (7413): 488–93، 2003، doi:10.1136/bmj.327.7413.488، PMC 188387، PMID 12946972.
- "Gastrointestinal symptoms and autism spectrum disorder: links and risks - a possible new overlap syndrome."، Pediatric Health Med Ther (Review)، 6: 153–166، 2015، doi:10.2147/PHMT.S85717، PMC 5683266، PMID 29388597.
- "The bowel and beyond: the enteric nervous system in neurological disorders"، Nat Rev Gastroenterol Hepatol (Review)، 13 (9): 517–28، سبتمبر 2016، doi:10.1038/nrgastro.2016.107، PMC 5005185، PMID 27435372.
- "Serotonin as a link between the gut-brain-microbiome axis in autism spectrum disorders."، Pharmacol Res (Review)، 132: 1–6، 2018، doi:10.1016/j.phrs.2018.03.020، PMC 6368356، PMID 29614380.
- "Autism"، Lancet، 374 (9701): 1627–38، 2009، doi:10.1016/S0140-6736(09)61376-3، PMC 2863325، PMID 19819542.
- "The emerging role of synaptic cell-adhesion pathways in the pathogenesis of autism spectrum disorders"، Trends Neurosci، 32 (7): 402–12، 2009، doi:10.1016/j.tins.2009.04.003، PMID 19541375، S2CID 8644511، مؤرشف من الأصل في 15 نوفمبر 2020.
- "Autism and brain development"، Cell، 135 (3): 396–400، 2008، doi:10.1016/j.cell.2008.10.015، PMC 2701104، PMID 18984148.
- "The teratology of autism"، Int J Dev Neurosci، 23 (2–3): 189–99، 2005، doi:10.1016/j.ijdevneu.2004.11.001، PMID 15749245، S2CID 17797266.
- "Autism: many genes, common pathways?"، Cell، 135 (3): 391–95، 2008، doi:10.1016/j.cell.2008.10.016، PMC 2756410، PMID 18984147.
- "The study of autism as a distributed disorder"، Ment Retard Dev Disabil Res Rev، 13 (1): 85–95، 2007، doi:10.1002/mrdd.20141، PMC 3315379، PMID 17326118.
- "Neuroanatomy of autism"، Trends Neurosci، 31 (3): 137–45، 2008، doi:10.1016/j.tins.2007.12.005، PMID 18258309، S2CID 18648870.
- "The neuropathology of autism"، Brain Pathol، 17 (4): 422–33، 2007، doi:10.1111/j.1750-3639.2007.00100.x، PMID 17919128، S2CID 6959302، مؤرشف من الأصل في 23 يناير 2021.
- "Advances in autism"، Annu Rev Med، 60: 367–80، 2009، doi:10.1146/annurev.med.60.053107.121225، PMC 3645857، PMID 19630577.
- "Mapping early brain development in autism"، Neuron، 56 (2): 399–413، 2007، doi:10.1016/j.neuron.2007.10.016، PMID 17964254، S2CID 10662307.
- "The neuropathology of autism: where do we stand?"، Neuropathol Appl Neurobiol، 34 (1): 4–11، 2008، doi:10.1111/j.1365-2990.2007.00872.x، PMID 17971078، S2CID 23551620.
- "Searching for ways out of the autism maze: genetic, epigenetic and environmental clues"، Trends Neurosci، 29 (7): 349–58، 2006، doi:10.1016/j.tins.2006.05.010، PMID 16808981، S2CID 26722022.
- "Neuroligins and neurexins link synaptic function to cognitive disease"، Nature، 455 (7215): 903–11، 2008، Bibcode:2008Natur.455..903S، doi:10.1038/nature07456، PMC 2673233، PMID 18923512.
- "The autistic neuron: troubled translation?"، Cell، 135 (3): 401–06، 2008، doi:10.1016/j.cell.2008.10.017، PMID 18984149، S2CID 619383.
- "Fragile X: translation in action"، Neuropsychopharmacology، 33 (1): 84–7، 2008، doi:10.1038/sj.npp.1301610، PMC 4327813، PMID 17940551.
- "Convulsing toward the pathophysiology of autism"، Brain Dev، 31 (2): 95–103، 2009، doi:10.1016/j.braindev.2008.09.009، PMC 2734903، PMID 19006654.
- "Immune Dysregulation in Autism Spectrum Disorder"، Neurobiology of Autism، International Review of Neurobiology، ج. 113، 2013، ص. 269–302، doi:10.1016/B978-0-12-418700-9.00009-5، ISBN 9780124187009، PMID 24290389.
- "The role of immune dysfunction in the pathophysiology of autism"، Brain, Behavior, and Immunity، 26 (3): 383–92، أغسطس 2011، doi:10.1016/j.bbi.2011.08.007، PMC 3418145، PMID 21906670.
- "Evidence linking oxidative stress, mitochondrial dysfunction, and inflammation in the brain of individuals with autism"، Frontiers in Physiology، 5: 150، 2014، doi:10.3389/fphys.2014.00150، PMC 4001006، PMID 24795645.
- "Maternal infection and immune involvement in autism"، Trends in Molecular Medicine، 17 (7): 389–94، يوليو 2011، doi:10.1016/j.molmed.2011.03.001، PMC 3135697، PMID 21482187.
- "Autism risk factors: genes, environment, and gene-environment interactions"، Dialogues Clin Neurosci، 14 (3): 281–92، 2012، PMC 3513682، PMID 23226953.
- "The immune response in autism: a new frontier for autism research"، J Leukoc Biol، 80 (1): 1–15، 2006، CiteSeerX 10.1.1.329.777، doi:10.1189/jlb.1205707، PMID 16698940، S2CID 17531542، مؤرشف من الأصل في 05 أكتوبر 2006.
- "Maternal hospitalization with infection during pregnancy and risk of autism spectrum disorders"، Brain, Behavior, and Immunity، 44: 100–105، سبتمبر 2014، doi:10.1016/j.bbi.2014.09.001، PMC 4418173، PMID 25218900.
- "Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders"، Journal of Autism and Developmental Disorders، 40 (12): 1423–30، ديسمبر 2010، doi:10.1007/s10803-010-1006-y، PMID 20414802، S2CID 23471371.
- "Correction of fragile X syndrome in mice"، Neuron، 56 (6): 955–62، 2007، doi:10.1016/j.neuron.2007.12.001، PMC 2199268، PMID 18093519.
- "Update on autism: A review of 1300 reports published in 2008"، Epilepsy Behav، 16 (4): 569–89، 2009، doi:10.1016/j.yebeh.2009.09.023، PMID 19896907، S2CID 8013774.
- "Autism and metabolic diseases"، J Child Neurol، 23 (3): 307–14، 2008، doi:10.1177/0883073807308698، PMID 18079313، S2CID 30809774.
- MNS and autism:
- "Self–other relations in social development and autism: multiple roles for mirror neurons and other brain bases"، Autism Res، 1 (2): 73–90، 2008، doi:10.1002/aur.15، PMID 19360654، S2CID 15269399.
- "A mirror up to nature"، Curr Biol، 18 (1): R13–18، 2008، doi:10.1016/j.cub.2007.11.004، PMC 2517574، PMID 18177704، مؤرشف من الأصل في 24 يناير 2021.
- "The mirror neuron system and the consequences of its dysfunction"، Nature Reviews Neuroscience، 7 (12): 942–51، 2006، doi:10.1038/nrn2024، PMID 17115076، S2CID 9463011، مؤرشف من الأصل في 17 أكتوبر 2019.
- "Development and neurophysiology of mentalizing"، Philosophical Transactions of the Royal Society B، 358 (1431): 459–73، 2003، doi:10.1098/rstb.2002.1218، PMC 1693139، PMID 12689373.
- "Emulation and mimicry for social interaction: a theoretical approach to imitation in autism"، Q J Exp Psychol، 61 (1): 101–15، 2008، doi:10.1080/17470210701508798، PMID 18038342، S2CID 14569936، مؤرشف من الأصل في 17 أكتوبر 2019.
- "Functional brain correlates of social and nonsocial processes in autism spectrum disorders: an activation likelihood estimation meta-analysis"، Biol Psychiatry، 65 (1): 63–74، 2009، doi:10.1016/j.biopsych.2008.09.022، PMC 2993772، PMID 18996505.
- "Default-mode brain dysfunction in mental disorders: a systematic review"، Neurosci Biobehav Rev، 33 (3): 279–96، 2009، doi:10.1016/j.neubiorev.2008.09.002، PMID 18824195، S2CID 7175805، مؤرشف من الأصل في 27 مايو 2020.
- "Functional and anatomical cortical underconnectivity in autism: evidence from an FMRI study of an executive function task and corpus callosum morphometry"، Cereb Cortex، 17 (4): 951–61، 2007، doi:10.1093/cercor/bhl006، PMC 4500121، PMID 16772313، مؤرشف من الأصل في 07 يوليو 2010.
- "Neuropsychologic functioning in children with autism: further evidence for disordered complex information-processing"، Child Neuropsychol، 12 (4–5): 279–98، 2006، doi:10.1080/09297040600681190، PMC 1803025، PMID 16911973.
- "Resting state cortical connectivity reflected in EEG coherence in individuals with autism"، Biol Psychiatry، 62 (3): 270–73، 2007، doi:10.1016/j.biopsych.2006.11.012، PMC 2001237، PMID 17336944.
- "The new neurobiology of autism: cortex, connectivity, and neuronal organization"، Arch Neurol، 64 (7): 945–50، 2007، doi:10.1001/archneur.64.7.945، PMC 2597785، PMID 17620483.
- "Event related potentials in the understanding of autism spectrum disorders: an analytical review"، J Autism Dev Disord، 39 (3): 495–510، 2009، doi:10.1007/s10803-008-0652-9، PMC 4422389، PMID 18850262.
- "Electrophysiological signatures: magnetoencephalographic studies of the neural correlates of language impairment in autism spectrum disorders"، Int J Psychophysiol، 68 (2): 149–60، 2008، doi:10.1016/j.ijpsycho.2008.01.012، PMC 2397446، PMID 18336941.
- "Evolution in health and medicine Sackler colloquium: Comparative genomics of autism and schizophrenia"، Proceedings of the National Academy of Sciences of the United States of America، 107 (Suppl 1): 1736–41، 2010، Bibcode:2010PNAS..107.1736C، doi:10.1073/pnas.0906080106، PMC 2868282، PMID 19955444.
- "Characteristics of Brains in Autism Spectrum Disorder: Structure, Function and Connectivity across the Lifespan"، Exp Neurobiol (Review)، 24 (4): 273–84، ديسمبر 2015، doi:10.5607/en.2015.24.4.273، PMC 4688328، PMID 26713076.
- "Autism: the empathizing–systemizing (E-S) theory" (PDF)، Annals of the New York Academy of Sciences، 1156 (1): 68–80، 2009، Bibcode:2009NYASA1156...68B، doi:10.1111/j.1749-6632.2009.04467.x، PMID 19338503، S2CID 1440395، مؤرشف من الأصل (PDF) في 28 نوفمبر 2020.
- "Goals, intentions and mental states: challenges for theories of autism"، J Child Psychol Psychiatry، 50 (8): 881–92، 2009، CiteSeerX 10.1.1.621.6275، doi:10.1111/j.1469-7610.2009.02098.x، PMID 19508497.
- "Understanding executive control in autism spectrum disorders in the lab and in the real world"، Neuropsychol Rev، 18 (4): 320–38، 2008، doi:10.1007/s11065-008-9077-7، PMC 2856078، PMID 18956239.
- "Neurodevelopment and executive function in autism"، Dev Psychopathol، 20 (4): 1103–32، 2008، doi:10.1017/S0954579408000527، PMID 18838033.
- "Executive dysfunction in autism"، Trends Cogn Sci، 8 (1): 26–32، 2004، doi:10.1016/j.dr.2004.01.001، PMID 14697400، مؤرشف من الأصل في 23 يناير 2021.
- "Autism from developmental and neuropsychological perspectives"، Annual Review of Clinical Psychology، 2: 327–55، 2006، doi:10.1146/annurev.clinpsy.2.022305.095210، PMID 17716073.
- "The weak coherence account: detail-focused cognitive style in autism spectrum disorders"، Journal of Autism and Developmental Disorders، 36 (1): 5–25، يناير 2006، doi:10.1007/s10803-005-0039-0، PMID 16450045، S2CID 14999943، مؤرشف من الأصل في 17 أكتوبر 2019.
- "Enhanced perceptual functioning in autism: an update, and eight principles of autistic perception"، Journal of Autism and Developmental Disorders، 36 (1): 27–43، يناير 2006، doi:10.1007/s10803-005-0040-7، PMID 16453071، S2CID 327253.
- "Attention, monotropism and the diagnostic criteria for autism" (PDF)، Autism، 9 (2): 139–56، مايو 2005، doi:10.1177/1362361305051398، PMID 15857859، S2CID 6476917، مؤرشف من الأصل (PDF) في 19 مايو 2018، اطلع عليه بتاريخ 18 مارس 2018.
- "Time to give up on a single explanation for autism"، Nature Neuroscience، 9 (10): 1218–20، 2006، doi:10.1038/nn1770، PMID 17001340.
- "Cognitive theories of autism" (PDF)، Dev Rev، 27 (2): 224–60، 2007، doi:10.1016/j.dr.2007.02.001، مؤرشف من الأصل (PDF) في 6 ديسمبر 2018.
 بوابة علم النفس
بوابة علم النفس بوابة طب
بوابة طب