إنتئين
الإنتئين هو جزء من البروتين لديه القدرة على إزالة نفسه والانضمام مرة أخرى للأجزاء الباقية (الإكستينات) برابطة ببتيد. كما تمت تسمية الإنتئينات بإنترونات البروتين أيضاً.[1]
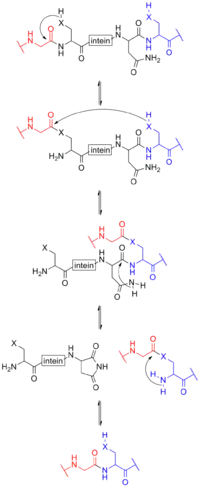
تحتوي معظم الإنتئينات على نطاق الإندونكلياز الذي يلعب دوراً في تكاثر الإنتئين. وفي الواقع فإن كثيراً من الجينات لديها قطع غير مترابطة بشفرة إنتئينية متواجدة في مواضع متباينة. ولأجل ذلك وغيرها من الأسباب، يسمى الإنتئين أحياناً أو بشكل أدق قِطع الجين الرامزة للإنتئين بالعناصر الجينية الأنانية، لكنه من الأدق تسميتها بالطفيلية. ويكمن الفرق في أن الجينات الأنانية أنانية بقدر تنافسها مع الجينات الأخرى أو الأليلات ولكنها تؤدي وظيفة، بينما تعد الجينات الطفيلية عديمة الفائدة.
يحدث تظفير البروتين بواسطة الإنتئين (بالإنجليزية: Intein-mediated protein splicing) بعد أن يُترجَم مرسال الحمض الريبي النووي (الرنا المرسال) إلى بروتين. ويحتوي هذا البروتين الطليعي على ثلاث قطع، إكستين إن متبوعاً بالإنتئين متبوعاً بإكستين سي. وبعد أن تتم عملية التظفير، يسمى الناتج بالإكستين أيضاً.
تم اكتشاف أول إنتئين في 1988 من خلال مقارنة التسلسل بين فطر النوروسبورا [2] وأتبيز الجزر الفجوي [3] (الذي لا يحوي إنتئين) والجين المتناظر في الخميرة (المحتوي على إنتئين) الذي وُصف في البداية بأنه ناقل مفترض لأيون الكالسيوم.[4] وفي 1990 برهن هيراتا وغيره [5] بأن التسلسل الزائد في جين الخميرة استحال إلى رنا مرسال وأزال نفسه من البروتين المضيف بعدما تمت عملية الترجمة فقط. منذ ذلك الحين تم العثور على الإنتئين في جميع مجالات الحياة الثلاثة (الخلايا حقيقة النواة والبكتريا والعتائق) وفي الفيروسات أيضاً. وتم استعراض المعارف بشأن الحالة التطورية للإنتئينات والعناصر المرتبطة بها في جوجارتن وهيلاريو عام 2006. وتعتبر آلية أثر التظفير تماثلاً طبيعي الحدوث لتقنية البروتينات ذات الحجم المتوسط المُولدة كيميائياً والمسماة بالربط الكيميائي الطبيعي والتي تم تطويرها في نفس الوقت الذي تم فيه اكتشاف الإنتئينات.
الإنتئينات في التكنولوجيا الحيوية
تلعب الإنتئينات دوراً هاماً في التكنولوجيا الحيوية لما لها من فعالية عالية في تظفير البروتين. وقد تم التعرف على أكثر من 200 إنتئين حتى الآن تتراوح أحجامها بين 100-800 حمض أميني. وصممت الإنتئينات لأجل تطبيقات معينة مثل تخليق البروتين [6] والتوسيم الانتقائي لقطع البروتين وهو أمر مفيد في دراسات الرنين النووي المغناطيسي للبروتينات الكبيرة.[7]
وقد يكون التثبيط الدوائي لإزالة الإنتئين أداة نافعة لتطوير الأدوية، فالبروتين المحتوي على إنتئين لن يواصل القيام بوظيفته المعتادة إذا لم يتم إزالة الإنتئين حيث أن تركيبه سيتعطل. كما قيل بأن الإنتئينات قد تكون مفيدة لتحقيق المصطلح الألوتوبيكي (بالإنجليزية: Allotopic expression) لبضع من البروتينات الكارهة للماء بشكل كبير والتي يرمز لها بجينيوم الميتوكندريا، كما في العلاج الجيني على سبيل المثال (دي غراي 2000). وتعتبر الطبيعة الكارهة للماء لهذه البروتينات عقبة أمام استيرادها في الميتوكندريا. وتبعاً لذلك، فإن إدراج إنتين ليس ذا طبيعة كارهة للماء قد يسمح لهذا الاستيراد بأن يستمر. وإزالة الإنتئين بعد استيراده سيعيد البروتين إلى النمط البري.
اصطلاحات تسمية الإنتئين
يرجع القسم الأول من اسم الإنتئين إلى الاسم العلمي للكائن الذي وُجد فيه، أما القسم الثاني فعائد إلى اسم الجين الملازم أو الإكستين. فعلى سبيل المثال، يُسمى الإنتئين الذي تم العثور عليه في الثيرموبلازما أسيدوفيلوم والمنتمي للوحدة أي في الأتيبيز الفجوي VMA بـ تاك في إم أي 'Tac VMA'. في العادة وكما في المثال، تكفي ثلاثة حروف لتعريف الكائن مع وجود أشكال مختلفة. فمثلاً، يمكن إضافة أحرف إضافية للإشارة إلى سلالة. وفي حال ترميز أكثر من إنتئين لدى الجين الملازم تُمنح الإنتئينات لاحقة عددية بدئاً من 5' حتى 3' أو على حسب ترتيب معرفتهم، مثل «مسم دنا بي-1»،"Msm dnaB-1". تُسمى قطعة الجين التي ترمز للإنتئين بنفس اسم الإنتئين، ومنعاً للبس يتم الرمز للإنتئين الصحيح بحروف كبيرة (e.g. Pfu RIR1-1), بينما يُرمز لقطعة الجين الملازم بالأحرف المائلة.
الإنتئينات الكاملة والصغيرة
يمكن للإنتئينات أن تحوي على نطاق جين توجيه الإندونكلياز (HEG) علاوة على نطاقات التظفير. ويعد هذا النطاق مسئولا عن انتشار الإنتئين عن طريق فصله الحمض النووي لأليل حر من الإنتئين في الكروموسومات المتناددة، مؤدياً إلى نظام ترميم قطوعات الدنا ثنائية الطاق (DSBR) التي ترمم القطع بدورها، ناسخةً الإنتئين إلى موقع إنتئين حر سابق. ولا يعد نطاق جين توجيه الإندونكلياز ضرورياً لتظفير الإنتئين، لذا قد يُفقد مكوناً إنتئينات صغيرة. وأثبتت بعض الدراسات طبيعة الإنتئينات المعيارية التي تتحقق بإضافة أو إزالة نطاقات جين توجيه الإندونكلياز وتحديد نشاط التركيب الجديد.
الإنتئين المنقسم
يَنتج إنتئين البروتين الطليعي في بعض الأحيان من جينين. وفي هذا الحالة يُقال بأن الإنتئين إنتئين منقسم. فعلى سبيل المثال، في الزراقم يُرمز للوحدة الفرعية ألفا المحفزة لبلمرة الدنا الثالث، DnaE، بجينين مختلفين هما dnaE-n و dnaE-c. ويتكون الـdnaE-n من سلسلة إكستين إن متبوعاً بسلسلة إنتئين ذات 123حمض أميني، في حين يحتوي الـdnaE-c سلسلة إنتئين ذات 36 حمض أميني متبوعة بسلسلة اكستين سي.
اقرأ أيضاً
- صراع الجينيوم الداخلي (Intragenomic conflict)
المراجع
- de Grey, Aubrey D. N. J. (2000)، "Mitochondrial gene therapy: an arena for the biomedical use of inteins"، Trends Biotechnol، 18 (9): 394–399، doi:10.1016/S0167-7799(00)01476-1، ISSN 0167-7799، PMID 10942964، مؤرشف من الأصل في 10 سبتمبر 2020، اطلع عليه بتاريخ 30 نوفمبر 2008.
- Gogarten, J Peter (2006)، "Inteins, introns, and homing endonucleases: recent revelations about the life cycle of parasitic genetic elements"، BMC Evol Biol، 6 (1): 94، doi:10.1186/1471-2148-6-94، ISSN 1471-2148، PMC 1654191، PMID 17101053، مؤرشف من الأصل في 20 يونيو 2015، اطلع عليه بتاريخ 30 نوفمبر 2008.
- Anraku Y, Mizutani R, Satow Y (2005)، "Protein splicing: its discovery and structural insight into novel chemical mechanisms"، IUBMB Life، 57 (8): 563–74، doi:10.1080/15216540500215499، PMID 16118114.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Bowman EJ, Tenney K, Bowman BJ Isolation of genes encoding the Neurospora vacuolar ATPase. Analysis of vma-1 encoding the 67-kDa subunit reveals homology to other ATPases. J Biol Chem. 1988 Oct 5;263(28):13994-4001.
- Zimniak L, Dittrich P, Gogarten JP, Kibak H, Taiz L.The cDNA sequence of the 69-kDa subunit of the carrot vacuolar H+-ATPase. Homology to the beta-chain of F0F1-ATPases. J Biol Chem. 1988 Jul 5;263(19):9102-12
- Shih CK, Wagner R, Feinstein S, Kanik-Ennulat C, Neff N. A dominant trifluoperazine resistance gene from Saccharomyces cerevisiae has homology with F0F1 ATP synthase and confers calcium-sensitive growth. Mol Cell Biol. 1988 Aug;8(8):3094-103
- Hirata R, Ohsumk Y, Nakano A, Kawasaki H, Suzuki K, Anraku Y. Molecular structure of a gene, VMA1, encoding the catalytic subunit of H(+)-translocating adenosine triphosphatase from vacuolar membranes of Saccharomyces cerevisiae. J Biol Chem. 1990 Apr 25;265(12):6726-33.
- Schwarzer D, Cole PA (2005)، "Protein semisynthesis and expressed protein ligation: chasing a protein's tail"، Curr Opin Chem Biol، 9 (6): 561–9، doi:10.1016/j.cbpa.2005.09.018، PMID 16226484.
- Muralidharan V, Muir TW (2006)، "Protein ligation: an enabling technology for the biophysical analysis of proteins"، Nat. Methods، 3 (6): 429–38، doi:10.1038/nmeth886، PMID 16721376.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء