الحلزون المزدوج للحمض النووي
في علم الأحياء الجزيئي ، يشير مصطلح اللولب المزدوج [1] إلى البنية المكونة من جزيئات الأحماض النووية المزدوجة الشريطية مثل الحمض النووي .تنشأ بنية اللولب المزدوج لمعقد الحمض النووي نتيجة للمستوى الثانوي الذي تمتلكه ، وهو مكون أساسي في تحديد هيكله الثالث . وقد دخل المصطلح إلى الثقافة الشعبية مع نشر مصطلح اللولبالمزدوج في عام 1968 : حساب شخصي لاكتشاف بنية الحمض النووي بواسطة جيمس واتسون
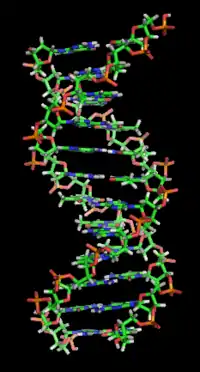
ويُحتجز الحمض النووي مزدوج الهيدروكسيد ثنائي الأحماض الحيوية في الحمض النووي معًا بواسطة النيوكليوتيدات التي تتكون منها القاعدة الثنائية [2] ويوجد في الحمض النووي بي الهيكل الحلزوني المزدوج الأكثر شيوعًا الموجود في الطبيعة ، يكون اللولب المزدوج يمينًا بحوالي 10 – 10.5 أزواج قاعدية لكل دور. [3] يحتوي التركيب الحلزوني المزدوج للحمض النووي على أخدود رئيسي وأخدود ثانوي . في في الحمض النووي ب، يكون الأخدود الرئيسي أوسع من الأخدود الصغير. نظرًا للاختلاف في عرض الأخدود الرئيسي والأخدود الصغير ، فإن العديد من البروتينات التي ترتبط بـ في الحمض النووي ب تفعل ذلك من خلال الأخدود الرئيسي الأوسع. [4]
التاريخ
تم نشر نموذج اللولب المزدوج لبنية الحمض النووي لأول مرة في مجلة الطبيعة من قبل جيمس و وفرانسيس كريك في عام 1953، (إحداثيات الاشعة السينية في عام 1954 بناءً على عمل روزاليند فرانكلين ، بناءً على عمل روزاليند فرانكلين، بما في ذلك صورة حيود الأشعة السينية الحيوية للحمض النووي "، من عام 1952، [5] تليها صورت الحمض النووي أكثر أوضح مع ريمون فرخ الوز ، [6] [7] موريس ويلكنز ، الكسندر ستوكس ، و هربرت ويلسون ، [8] وإقران قاعدة المعلومات الكيميائية والبيوكيميائية من قبل اروين تشارجاف . [9] [10] [11] [12] [13] [14] كان النموذج السابق عبارة عن حمض نووي ثلاثي الجديلة . [15]
إن إدراك بنية الحمض النووي عل أنها هي بنية الحلزون المزدوج لآلية الاقتران الأساسي الذي يتم من خلاله تخزين المعلومات الجينية ونسخها في الكائنات الحية ويعتبرمن أهم الاكتشافات العلمية بشكل كبير في القرن العشرين. وقد حصل كل من كريك و ويلسون و واتسون على ثلث جائزة نوبل لعام 1962 في علم وظائف الأعضاء أو الطب لمساهماتهم في الاكتشاف. [16]
تهجين الحمض النووي
التهجين هو عملية ربط أزواج القاعدة التكميلية لتكوين لولب مزدوج. الذوبان هو العملية التي يتم من خلالها كسر التفاعلات بين خيوط اللولبية المزدوج ، مما يفصل بين خيوط الحمض النووي. والواقع أن هذه الروابط ضعيفة، يسهل فصلها بالتسخين الناعم، أو الإنزيمات، أو القوة الميكانيكية. [17] يتم ذوبان المناطق الغنية ت و أ بسهولة أكبر من المناطق الغنية ج و د. بعض الخطوات الأساسية (الأزواج) عرضة أيضًا لذوبان الحمض النووي ، مثل أ ت و أ ج . [18] تنعكس هذه الميزات الميكانيكية من خلال استخدام متواليات مثل أ ت أ ت في بداية العديد من الجينات لمساعدة الحمض النووي الريبيبوليميراز في إذابة الحمض النووي للنسخ.
يعد فصل السلك عن طريق التسخين اللطيف ، كما هو مستخدم في تفاعل البوليميراز المتسلسل (PCR) ، أمرًا بسيطًا ، مما يوفر للجزيئات أقل من 10000 زوج قاعدي (10 أزواج كيلوباز ، أو 10 كيلو بايت). إن اقتران/تشابك جدائل(خيوط) الحمض النوي يجعل فصل الأجزاء الطويلة أمرًا صعبًا للغاية. وتتجنب الخلية هذه المشكلة بالسماح بإنزيمات انصهار الحمض النووي لديها (مساعدين) للعمل مع هيليكازات ) بالعمل بشكل متزامن مع الإيزوميراز العلوي ، والذي يمكنه تشق العمود الفقري للفوسفات في أحد الخيوط كيميائيًا بحيث يمكنه الدوران حول الآخر. تقوم الهيليكازات بفك الخيوط لتسهيل تقدم إنزيمات قراءة التسلسل مثل بوليميريز الحمض النووي.
هندسة زوج القاعدة
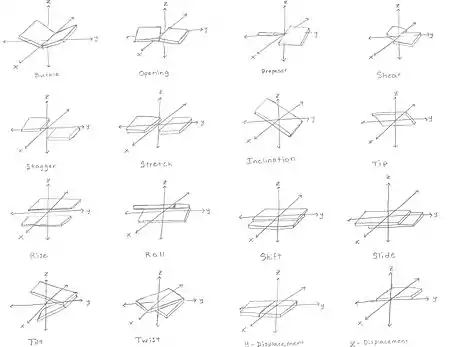
يمكن أن تتميز هندسة القاعدة أو خطوة الزوج الأساسي بـ 6 مميزات: وهي الإزاحة ، الانزلاق ، الارتفاع ، الإمالة ، اللف ، والتدوير.حيث تحدد هذه القيم موقع واتجاه كل زوج من القواعد أو القاعدة في جزيء حمض نووي بالنسبة إلى سلفه على طول محور اللولبة بشكل دقيق .. معًا ، يميزان التركيب الحلزوني للجزيء. في مناطق الحمض النووي أو الحمض النووي الريبي حيث يتم تعطيل البنية الطبيعية ، يمكن استخدام التغيير في هذه القيم لوصف هذا الاضطراب.
بالنسبة لكل زوج من أزواج القواعد، يتم اعتباره بالنسبة إلى سابقه، كما أن هناك هندسة لزوج القاعدة التالية التي يجب أخذها في الاعتبار:: [19] [20] [21]
- قص
- تمتد
- ترنح
- انبعاج
- المروحة : دوران قاعدة واحدة بالنسبة إلى الأخرى في نفس زوج القاعدة.
- افتتاح
- التحول : الإزاحة على طول محور في مستوى زوج القاعدة عموديًا على الأول ، موجهًا من الأخدود الصغير إلى الأخدود الرئيسي.
- الانزلاق : الإزاحة على طول محور في مستوى زوج القاعدة الموجه من حبلا إلى الآخر.
- الارتفاع : الإزاحة على طول محور اللولب.
- الإمالة : الدوران حول محور التحول.
- لفة : دوران حول محور الشريحة.
- الالتواء : الدوران حول محور الارتفاع.
- x الإزاحة
- ص الإزاحة
- ميل
- تلميح
- الملعب : الارتفاع لكل دورة كاملة للحلزون.
الصعود والالتواء يقومان بتحديد مدى الازدحام والميل من اللولب.. أما الإحداثيات الأخرى، على النقيض من ذلك، فيمكن أن تكون صفرًا. عادةً ما يكون الانزلاق والتحول صغيرًا في الحمض النووي ب، لكنهما كبيران في الحمض النووي أ-س. يجعل التدحرج والإمالة أزواج القاعدة المتتالية أقل توازيًا ، وعادة ما تكون صغيرة.
لاحظ أن "الميل" قد بتم استخدامه في كثير من الأحيان بشكل مختلف في الكتابات العلمية، مع الإشارة إلى انحراف محور زوج القاعدة الأول، بين الجديلين بشكل عمودي نادرة إلى المحور الحلزوني. هذا يتوافق مع الانزلاق بين سلسلة من أزواج القاعدة ، وفي الإحداثيات القائمة على الحلزون يسمى بشكل صحيح "الميل".
هندسة الحلزون
ويُعتقد أن ثلاثة تشكيلات من الحمض النووي موجودة في الطبيعة، الحمض النووي أ، الحمض النووي ب، الحمض النووي س. ويعتقد النموذج ب الذي وصفه جيمس واطسون وفرانسيس كريك على أنه يقوم بالسيطرة على الخلايا [22] التي يبلغ عرضها 23.7 وتمتد بمقدار 34 لكل 10 نقاط أساس من التسلسل. يقوم اللولب المزدوج بعمل دورة كاملة حول محوره كل 10.4-10.5 أزواج قاعدية في المحلول. هذا التردد من تطور (وهو ما يسمى أرض الملعب حلزونية) يعتمد إلى حد كبير على القوات التراص أن كل قاعدة تمارس على جيرانها في السلسلة. يحدد التكوين المطلق للقواعد اتجاه المنحنى الحلزوني لتشكل معين.
يختلفان الحمض النووي أ والحمض النووي س بشكل كبير في هندستهما وأبعادهما عن الحمض النووي ب، على الرغم من أنهما لا يزالان يشكلان هياكل حلزونية. كان اعتقاد العلماء لفترة طويلة ب أن الشكل لا يحدث إلا في عينات من الحمض النووي المجمد في المختبر، مثل تلك المستخدمة في التجارب البلورية ، وفي التزاوج الهجين لخيوط الحمض النووي والحمض الاميني، لكن جفاف الحمض النووي يحدث في الجسم الحي ، والحمض النووي أ هو من المعروف الآن أن لها وظائف بيولوجية . قد تتبنى أجزاء الحمض النووي التي قامت الخلايا ميثليتها لأغراض تنظيمية الهندسة س ، حيث تدور الخيوط حول المحور الحلزوني في الاتجاه المعاكس لـ الحمض النووي أ و الحمض النووي ب. هناك أيضًا دليل على أن مجمعات البروتين للحمض النووي تشكل هياكل الحمض النووي س.
ومن الممكن حدوث تشوهات أخرى؛ للأحماض النوية أ -ب-ج-ه، (الشكل التماوج من الاحماض النووية ب- س-ز-د) ، [23] ا ، [24] ال ، إلخ. تم وصفه حتى الآن. [25] في الواقع ، الأحرف في وكيو و يو و في و واي فقط هي اعتبارًا من 17 فبراير 2011 متاح لوصف أي بنية الحمض النووي جديدة قد تظهر في المستقبل. [26] [27] ومع ذلك ، فقد تم إنشاء معظم هذه الأشكال صناعياً ولم يتم ملاحظتها في الأنظمة البيولوجية التي تحدث بشكل طبيعي. هناك أيضًا أشكال الحمض النووي الثلاثية الجديلة وأشكال رباعية مثل مجموعة ال [إ4-بربلكس] و [إي-موتيف].
| سمة الهندسة | A- DNA | ب- DNA | Z-DNA |
|---|---|---|---|
| حاسة الحلزون | أيمن | أيمن | يساري |
| وحدة التكرار | 1 زوج | 1 زوج | 2 بي بي |
| التناوب / bp | 32.7 درجة | 34.3 درجة | 60 درجة / 2 |
| بي بي / بدوره | 11 | 10.5 | 12 |
| ميل bp إلى المحور | + 19 درجة | −1.2 درجة | −9 درجة |
| ارتفاع / بي بي على طول المحور | 2.3 Å (0.23 نانومتر) | 3.32 Å (0.332 نانومتر) | 3.8 Å (0.38 نانومتر) |
| الملعب / بدوره اللولب | 28.2 Å (2.82 نانومتر) | 33.2 Å (3.32 نانومتر) | 45.6 Å (4.56 نانومتر) |
| يعني تحريف المروحة | + 18 درجة | + 16 درجة | 0 درجة |
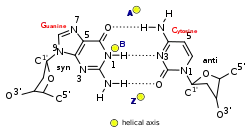
| زاوية الجليكوزيل | مضاد | مضاد | ج: مضاد ، </br> G: مزامنة |
| تجعد السكر | C3'- إندو | C2'- إندو | ج: C2'- إندو ، </br> G: C2'-exo |
| قطر الدائرة | 23 Å (2.3 نانومتر) | 20 Å (2.0 نانومتر) | 18 Å (1.8 نانومتر) |
الحزوز
تشكل الخيوط الحلزونية المزدوجة العمود الفقري للحمض النووي. يمكن العثور على حلزون مزدوج آخر عن طريق تتبع المسافات ، أو الأخاديد ، بين الخيوط. هذه الفراغات متاخمة للأزواج الأساسية وقد توفر موقع ربط . نظرًا لأن الخيوط لا تتقابل مباشرة مع بعضها البعض ، فإن حجم الأخاديد غير متساوٍ. أخدود واحد ، الأخدود الرئيسي ، هو 22 Å عرضًا والآخر ، الأخدود الصغير ، هو 12 واسع. [31] يعني ضيق الأخدود الصغير أن حواف القواعد يمكن الوصول إليها بشكل أكبر في الأخدود الرئيسي. ونتيجة لذلك ، فإن البروتينات مثل عوامل النسخ التي يمكن أن ترتبط بتسلسلات معينة في الحمض النووي مزدوج الشريطة عادةً ما تقوم بالاتصال بجوانب القواعد المكشوفة في الأخدود الرئيسي. [4] يختلف هذا الموقف في التوافقات غير العادية للحمض النووي داخل الخلية (انظر أدناه) ، ولكن يتم دائمًا تسمية الأخاديد الرئيسية والثانوية لتعكس الاختلافات في الحجم التي يمكن رؤيتها إذا تم التواء الحمض النووي مرة أخرى إلى شكل ب العادي.
الانحناء
تمتد
الالتفاف الفائق والطوبولوجيا
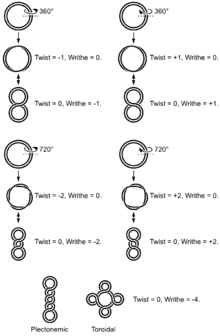
يلتف الشكل B للحلزون DNA بزاوية 360 درجة لكل 10.4-10.5 نقطة أساس في حالة عدم وجود إجهاد الالتواء. لكن العديد من العمليات البيولوجية الجزيئية يمكن أن تحفز إجهاد الالتواء. يُشار إلى مقطع DNA الذي يحتوي على التواء حلزوني زائد أو غير كاف ، على التوالي ، على أنه ملفوف بشكل إيجابي أو سلبي. عادةً ما يكون الحمض النووي في الجسم الحي ملفوفًا بشكل سلبي للغاية ، مما يسهل فك (ذوبان) الحلزون المزدوج المطلوب لنسخ الحمض النووي الريبي.
مراجع
- Kabai, Sándor (2007)، "Double Helix"، The Wolfram Demonstrations Project، مؤرشف من الأصل في 18 سبتمبر 2020.
- Alberts؛ وآخرون (1994)، The Molecular Biology of the Cell، New York: Garland Science، ISBN 978-0-8153-4105-5.
- Wang JC (1979)، "Helical repeat of DNA in solution"، PNAS، 76 (1): 200–203، Bibcode:1979PNAS...76..200W، doi:10.1073/pnas.76.1.200، PMID 284332.
- "Protein-DNA recognition"، Annu Rev Biochem، 53: 293–321، 1984، doi:10.1146/annurev.bi.53.070184.001453، PMID 6236744.
- "Secret of Photo 51"، Nova، PBS، مؤرشف من الأصل في 05 نوفمبر 2020.
- http://www.nature.com/nature/dna50/franklingosling.pdf نسخة محفوظة 2017-10-24 على موقع واي باك مشين.
- "The Structure of the DNA Molecule"، مؤرشف من الأصل في 21 يونيو 2012، اطلع عليه بتاريخ 30 أبريل 2010.
- "Molecular Structure of Deoxypentose Nucleic Acids" (PDF)، Nature، 171 (4356): 738–740، 1953، Bibcode:1953Natur.171..738W، doi:10.1038/171738a0، PMID 13054693، مؤرشف من الأصل (PDF) في 24 أكتوبر 2017.
- "On the deoxyribonucleic acid content of sea urchin gametes"، Experientia، 8 (4): 143–145، 1952، doi:10.1007/BF02170221، PMID 14945441.
- "Composition of the deoxypentose nucleic acids of four genera of sea-urchin"، J Biol Chem، 195 (1): 155–160، 1952، PMID 14938364.
- "The composition of the deoxyribonucleic acid of salmon sperm"، J Biol Chem، 192 (1): 223–230، 1951، PMID 14917668.
- Chargaff E (1951)، "Some recent studies on the composition and structure of nucleic acids"، J Cell Physiol Suppl، 38 (Suppl).
- "The separation and estimation of ribonucleotides in minute quantities"، J Biol Chem، 186 (1): 37–50، 1950، PMID 14778802.
- Chargaff E (1950)، "Chemical specificity of nucleic acids and mechanism of their enzymatic degradation"، Experientia، 6 (6): 201–209، doi:10.1007/BF02173653، PMID 15421335.
- "A proposed structure for the nucleic acids"، Proc Natl Acad Sci U S A، 39 (2): 84–97، فبراير 1953، Bibcode:1953PNAS...39...84P، doi:10.1073/pnas.39.2.84، PMID 16578429.
- "Nobel Prize - List of All Nobel Laureates"، مؤرشف من الأصل في 13 أغسطس 2018.
- "Predicting DNA duplex stability from the base sequence"، PNAS، 83 (11): 3746–3750، 1986، Bibcode:1986PNAS...83.3746B، doi:10.1073/pnas.83.11.3746، PMID 3459152.
- Owczarzy, Richard (28 أغسطس 2008)، "DNA melting temperature - How to calculate it?"، High-throughput DNA biophysics، owczarzy.net، مؤرشف من الأصل في 02 أغسطس 2020، اطلع عليه بتاريخ 02 أكتوبر 2008.
- Dickerson RE (1989)، "Definitions and nomenclature of nucleic acid structure components"، Nucleic Acids Res، 17 (5): 1797–1803، doi:10.1093/nar/17.5.1797، PMID 2928107.
- "Resolving the discrepancies among nucleic acid conformational analyses"، J Mol Biol، 285 (4): 1563–1575، 1999، doi:10.1006/jmbi.1998.2390، PMID 9917397.
- "A standard reference frame for the description of nucleic acid base-pair geometry"، J Mol Biol، 313 (1): 229–237، 2001، doi:10.1006/jmbi.2001.4987، PMID 11601858.
- Richmond؛ Davey, CA؛ وآخرون (2003)، "The structure of DNA in the nucleosome core"، Nature، 423 (6936): 145–150، Bibcode:2003Natur.423..145R، doi:10.1038/nature01595، PMID 12736678.
- "Application of L-DNA as a molecular tag"، Nucleic Acids Symp Ser (Oxf)، 49 (1): 261–262، 2005، doi:10.1093/nass/49.1.261، PMID 17150733.
- "Stretched and overwound DNA forms a Pauling-like structure with exposed bases"، PNAS، 95 (24): 14152–14157، 1998، Bibcode:1998PNAS...9514152A، doi:10.1073/pnas.95.24.14152، PMID 9826669.
- List of 55 fiber structures نسخة محفوظة 2007-05-26 على موقع واي باك مشين.
- Bansal M (2003)، "DNA structure: Revisiting the Watson-Crick double helix"، Current Science، 85 (11): 1556–1563.
- "A glossary of DNA structures from A to Z"، Acta Crystallogr D، 59 (4): 620–626، 2003، doi:10.1107/S0907444903003251، PMID 12657780.
- "The chemistry and biology of left-handed Z-DNA"، Annual Review of Biochemistry، 53: 791–846، 1984، doi:10.1146/annurev.bi.53.070184.004043، PMID 6383204.
- Sinden, Richard R (15 يناير 1994)، DNA structure and function (ط. 1st)، Academic Press، ص. 398، ISBN 0-12-645750-6.
- Ho PS (27 سبتمبر 1994)، "The non-B-DNA structure of d(CA/TG)n does not differ from that of Z-DNA"، Proc Natl Acad Sci USA، 91 (20): 9549–9553، Bibcode:1994PNAS...91.9549H، doi:10.1073/pnas.91.20.9549، PMID 7937803.
- "Crystal structure analysis of a complete turn of B-DNA"، Nature، 287 (5784): 755–8، 1980، Bibcode:1980Natur.287..755W، doi:10.1038/287755a0، PMID 7432492.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء