نظام غشائي داخلي
النظام الغشائي الداخلي[1] أو جهاز الغشاء الداخلي[2] (بالإنجليزية: Endomembrane system) هو نظامٌ مكونٌ من أغشيةٍ مُعلقةٍ في سيتوبلازم حقيقيات النوى، وتقسمُ هذه الأغشية الخلية إلى حجراتٍ وظيفية وبنيوية أو عضيات. تتضمن عضيات النظام الغشائي الداخلي في حقيقيات النوى: الغلاف النووي، والشبكة الإندوبلازمية، وجهاز غولجي، والأجسام الحالة، والحويصلات، والجُسيمات الداخلية، والغشاء البلازمي الخلوي وغيرها. يُعَرفُ النظام الغشائي الداخلي بشكلٍ دقيقٍ على أنهُ مجموعةٌ من الأغشية التي تُشكلُ وحدةً وظيفية وتطورية، والتي إما أن ترتبط مباشرةً أو تتبادل المواد بالحويصلات الناقلة.[3] يجب الإشارة أنَّ النظام الغشائي الداخلي لا يتضمن أغشية البلاستيدات الخضراء أو الميتوكندريا، ولكنهُ رُبما يكون قد تطورَ من الميتوكندريا.
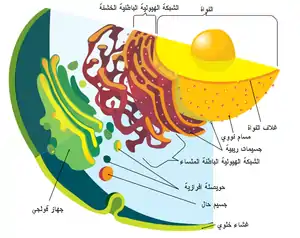
يحتوي الغلاف النووي على طبقةٍ مزدوجة من الليبيد والتي تُطوِقُ محتويات النواة.[4] الشبكة الإندوبلازمية هي عضيةٌ غشائية ناقلة ومُخلِّقة تمتدُ إلى السيتوبلازم في الخلايا النباتية والحيوانية.[5] جهاز غولجي هو سلسلةٌ من حجراتٍ مُتعددة والتي تُجمع فيها الجزئيات لنقلها إلى مكوناتِ الخلية الأخرى أو لإفرازها من الخلية.[6] تُوجد الفجوات العصارية في الخلايا الحيوانية والنباتية، ولكنها أكبر حجمًا في الخلايا النباتية، وهي مسؤولةٌ عن الحفاظ على شكل وهيكل الخلية، بالإضافة إلى تخزين مُخلفاتها،[7] أما الحويصلات فتكون صغيرةً نسبيًا، وهي كيسٍ مُغلقٍ بغشاء، يعمل على تخزين ونقلِ المواد.[8] الغشاء الخلوي هو حاجزٌ محصنٌ للخلية، حيثُ يُنظمُ ماذا يدخل ويخرج منها.[9] هُناك عُضية تعرف باسم الجسم المدبب حيثُ تتواجد فقط في الفطر، وترتبط مع نمو الطرف الخيطي.[10]
نادرًا ما تتواجدُ الأغشية الداخلية في بدائيات النوى، وذلك على الرغم من أنَّ العديد من بكتيريا التركيب الضوئي تحتوي على غشاءٍ بلازمي مطويٍ للغاية، ومعظم سيتوبلازم الخلية مليءٌ بطبقاتٍ من الأغشية المُجمعة للضوء،[11] وقد تشكلُ هذه الأغشية هيكلًا مُغلفًا يُسمى الجسم الأخضر، ويحدث هذا في بكتيريا الخَضْرَبيَّات.[12]
ترتبطُ العضُيات في النظام الغشائي الداخلي عبر الاتصال المُباشر أو عبر انتقالِ أجزاءٍ غشائية كالحويصلات، ولكن على الرغم من وجود هذه العلافة، إلا أنَّ الأغشية المتنوعة لا تتطابق في الوظيفة والبُنية الهيكلية. لا يتمُ إصلاح ثَخَانَةُ الأغشية وتركيبها الجزيئي وسُلوكها الأيضي، حيثُ أنها قد تتعدل أكثر من مرة خلال حياتها. من السمات المُوحدة والتي تتقاسمها جميع الأغشية هو وجود الطبقة المزدوجة من الليبيد، مع بروتيناتٍ ترتبط بأيٍ من الجانبين أو تعبرُ من خلالها.[13]
التاريخ
تُخَلق مُعظم الليبيدات في الخميرة إما على شكل جزئيات ليبيدية في الشبكة الإندوبلازمية أو في الميتوكندريون، كما أنَّ جزءً بسيطًا يُخلق في الغشاء البلازمي أو الغلاف النووي، وقد لا يُخلق فيها.[14][15] يبدأُ التخليق الحيوي للشحميات السفينجولية في الشبكة الإندوبلازمية، ولكنهُ يكتمل في جهاز غولجي.[16] نفس الأمر يحدث في الثدييات، مع وجود فرق بسيط في الخطوات الأولى القليلة للتخليق الحيوي لشحميات الإيثر والذي يحدث في البيروكسيسومات،[17] وبناءً على هذا، فإنَّ الأغشية المُختلفة التي تُغلف العضيات تحت الخلوية الأُخرى يجب أن تُبنى عبر نقل الليبيدات من مواقع التخليق هذه،[18] ولكن على الرغم أنهُ من الواضح أنَّ نقل الليبيدات يُعتبر عملًا أساسيًا في التخليق الحيوي للعضيات، إلا أنَّ آلية نقل الليبيدات عبر الخلايا غير مفهومة بشكلٍ جيد حتى الآن.[19]
يعتبر موريه ومولينهاور أولَ من اقترحَ بأنَّ الأغشية الخلوية تُشكلُ نظامًا واحدًا يتبادل المواد بين مكوناته، وكان ذلك في عام 1974،[20] حيثُ قدمَ هذا الاقتراح كطريقةٍ لتفسير كيف تتجمع الأغشية الليبيدية المُختلفة في الخلية، حيثُ تتجمع عبر تدفق الليبيدات من مواقع تخليق الليبيدات.[21] كانت فكرة تدفق الليبيدات عبر نظامٍ مستمر من الأغشية والحويصلات بديلةً لفكرة أنَّ الأغشية المختلفة هي كيانات مستقلة تكونت من نقل المكونات الليبيدية الحُرة مثل الأحماض الدهنية والستيرولات، عبر السيتوسول. الأهم من ذلك، أنَّ نقل الليبيدات عبر السيتوسول وتدفق الليبييدات المستمر عبر نظامٍ غشائي داخلي لا يعتبران عمليتين متعارضتين، وقد يحدث كلاهما في الخلايا.[18]
المكونات
الغلاف النووي

يُحيطُ الغلاف النووي بالنواة، حيثُ يفصل محتوياتها عن السيتوبلازم. يتكون الغلاف النووي من طبقتين من الأغشية، كلٌ منهما عبارة عن طبقة ثنائية الليبيد مع بروتيناتٍ مرتبطة.[22] الغلاف النووي الخارجي مُستمرٌ مع غلاف الشبكة الإندوبلازمية الخشنة، حيثُ يتميز هذا الجزء منه بوجود ريبوسوماتٍ مرتبطة بالسطح، كما يستمرُ الغلاف الخارجي مع الغلاف النووي الداخلي، حيثُ أنَّ الطبقتين تندمجان معًا عند ثقوبٍ صغيرة جدًا تُسمى المسامات النووية، وهي مساماتٌ تخرقُ الغلاف النووي، ويبلغ قطرها حوالي 120 نانومتر، كما تسمحُ بمرور الجزئيات بين النواة والسيتوبلازم، حيثُ تسمح بمرور جزيئاتٍ محددة ولا تسمح بمرور غيرها.[23] نظرًا لوجود هذه المسامات في مناطق مزدحمة بالجزيئات، فإنها تلعب دورًا مهمًا في فيزيولوجيا الخلايا. يُسمى الحيز بين الغلاف النووي الخارجي والداخلي بالحيز حول النواة، والذي يندمجُ مع تجويف الشبكة الإندوبلازمية الخشنة.
يتحددُ هيكلُ الغلاف النووي عبر شبكةٍ من الخيوط المتوسطة (خيوط بروتينية)، وتترتبُ هذه الشبكة بطانةٍ مشابهةٍ لشبكةٍ تسمى الصفيحة النووية، والتي ترتبط مع الكروماتينات والبروتينات الغشائية المتكاملة والمحتويات الأخرى للنواة، ويكون هذا على طول السطح الداخلي للنواة. يُعتقد بأنَّ الصفيحة النووية تُساعد في وصولِ المواد داخل النواة إلى المسامِ النووية، كما يعتقدُ أنها تساعدُ أيضًا في تفكك الغلاف النووي خلال الانقسام المتساوي، وفي إعادة تجميعه بعد نهاية العملية.[4]
تتميزُ المسامُ النووية بكفاءةٍ عاليةٍ في السماح بالمرور الانتقائي للمواد من وإلى النواة؛ وذلك لأنَّ الغلاف النووي يمتلكُ قدرًا كبيرًا من الازدحام، حيثُ يجبُ أن ينتقل الحمض النووي الريبوزي (RNA) والوحدات الفرعية للريبوسومات بشكلٍ متواصل من النواة إلى السيتوبلازم، كما أنهُ يجب السماحُ بإدخال بعض المواد الأساسية لنشاط النواة من السيتوبلازم، والتي تتضمن الهستونات والبروتينات المُنظمة للجين والحمض النووي الريبوزي منقوص الأكسجين (DNA) وبوليميرازات الحمض النووي الريبوزي وغيرها من المواد الأساسية الأخرى. يحتوي الغلاف النووي في الخلية الثديية النموذجية على 3000-4000 مجمعاتٍ مسامية. إذا كانت الخلية تُخلقُ الحمض النووي الريبوزي منقوص الأكسجين (DNA)، فإنَّ كل مجمعٍ مسامي يجب أن ينقل حوالي 100 جزيء هستون بالدقيقة. إذا كانت الخلية تنمو بسرعةٍ، فإنَّ كل مجمعٍ مسامي يجب أن ينقل أيضًا حوالي 6 وحداتٍ فرعية ريبوسومية كبيرة وصغيرة مجمعةٌ حديثًا في الدقيقة، حيثُ ينقلها من النواة إلى السيتوسول، ويتم استخدامها هُناك في تَخليق البروتينات.[24]
الشبكة الإندوبلازمية

الشكبة الإندوبلازمية (ER) هي عضية غشائية ناقلة ومُخلِّقة وتعتبر امتدادا للغشاء النووي. أكثر من نصف المجموع الكلي للأغشية في خلايا حقيقيات النوى عبارة عن شبكة إندوبلازمية. تتكون الشبكة الإندوبلازمية من أكياس مسطحة ونبيبات متفرعة يُعتقد أنها مترابطة، لكي يشكل غشاء الشبكة الإندوبلازمية صحيفة مستمرة تحيط بفراغ داخلي وحيد، يسمى هذا الفراغ الملفف بحدة لمعة الشبكة الإندوبلازمية، ويشار إليه كذلك بفراغ صهريج الشبكة الإندوبلازمية. تشغل اللمعة حوالي عشرة بالمئة من الحجم الإجمالي للخلية. يسمح غشاء الشبكة الإندوبلازمية بالنقل الاختياري للجزيئات بين اللمعة والسيتوبلازم، وبما أنه مرتبط بالغلاف النووي فذلك يوفر قناةً بين النواة والسيتوبلازم.[25]
للشبكة الإندوبلازمية دورٌ محوريٌ في إنتاج، معالجة، ونقل المركبات الكيميائية الحيوية لاستخدامها داخل وخارج الخية. غشائها هو موقعُ تخليق جميع البروتينات عبر الغشائية والليبيدات لمعظم عضيات الخلية، بما في ذلك الشبكة الإندوبلازمية نفسها، جهاز غولجي، اليحلولات، الدُخْلولات، المتقدرة، البيروكسيات، الحويصلات الإفرازية، الغشاء الخلوي. فضلا عن ذلك، معظم البروتينات التي ستغادر الخلية بالإضافة إلى تلك التي ستستقر في لمعة الشبكة الإندوبلازمية وجهاز غولجي واليحلولات يتم نقلها في البداية إلى الشبكة الإندوبلازمية. وعليه، فإن تواجُد العديد من البروتينات في الفراغ الصهريجي للمعةِ الشبكة الإندوبلازمية لايكون إلا مؤقتا، إذ أنها تمر من هنالك في طريقها نحو مواقع أخرى. تبقى بعض البروتينات بشكل دائم في لمعة الشبكة وتعرف باسم بروتينات الشبكة الإندوبلازمية المقيمة، وهي بروتينات خاصة تملك إشارة استبقاء مميزة مكونة من تسلسلٍ محددٍ من الأحماض الأمينية الذي يعمل على بقائها في الشبكة. من الأمثلة على أحد بروتينات الشبكة الإندوبلازمية المقيمة المهمة هو بروتين شابرون المعروف باسم البروتين الرابط للجلوبين المناعي (BiP) والذي يتعرف على البروتينات الأخرى التي تم تركيبها أو معالجتها بشكل خاطئ ويمنع إرسالها إلى مواقعها النهائية.[26]
تعمل الشبكة الإندوبلازمية على فرزِ بروتيناتٍ مصاحبٍ للترجمة. عديد الببتيد الذي يحتوي على تسلسل إشارة الشبكة الإندوبلازمية (ببتيد إشعاري) يتم التعرف عليه من قبل بروتين متعرف على الإشارة (SRP) والذي يوقف تخليق البروتين وينقل عديد الببتيد إلى غشاء الشبكة الإندوبلازمية ويحرره، يدخل عديد الببتيد إلى لمعة الشبكة عبر الموقع البروتيني Sec61[27][28] ثم تُستأنف عملية الترجمة.[29]
توجد منطقتان متميزتان -لكنهما متصلتان- من الشبكة الإندوبلازمية، تختلفان في البنية والوظيفة: الشبكة الإندوبلازمية الملساء والشبكة الإندوبلازمية الخشنة. سميت الشبكة الخشنة كذلك لأن سطحها السيتوبلازمي مغطى بالريبوسومات، حيث يعطيها ذلك منظرا مليئا بالنتوءات حين تُرى عبر المجهر الإلكتروني، أما الشبكة الملساء فتبدو ملساء لانعدام الريبوسومات في سطحها السيتوبلازمي.[30]
وظائف الشبكة الإندوبلازمية الملساء
في معظم الخلايا، تكون مناطق الشبكة الإندوبلازمية الملساء (SER) ملساء جزئيًا وخشنة جزئيًا أيضًا، ويطلق عليها في بعض الأحيان اسم الشبكة الإندوبلازمية الانتقالية لأنها تحتوي مواقع خروجٍ تتبرعم منها حويصلات ناقلة، حاملة لبروتينات حديثة التخليق تمضي نحو جهاز غولجي. في بعض الخلايا المتخصصة تكون الشبكة الملساء وافرة ولها وظائف إضافية. تعمل الشبكة الملساء في هذه الخلايا المتخصصة في عمليات أيضية متنوعة تشمل: تخليق الليبيدات، أيض الكربوهيدرات وإزالة سمية الأدوية والسموم. إنزيمات الشبكة الملساء أساسية لتخليق اللبيدات بما في ذلك الزيوت، اللبيدات الفوسفورية والستيرويدات. هرمونات الجنس لدى الفقاريات وهرمونات الستيرويد المُفرزة بواسطة الغدد الكظرية هي من بين الستيرويدات التي تنتجها الشبكة الملساء في الخلايا الحيوانية. الخلايا التي تُخلِّق هذه الهرمونات غنية بالشبكة الإندوبلازمية الملساء.[25][30]
خلايا الكبد مثال آخر على الخلايا المتخصصة التي تحتوي على شبكة ملساء وفيرة. توفر هذه الخلايا مثالا على دور الشبكة الملساء في أيض الكربوهيدرات، حيث تخزن خلايا الكبد الكربوهيدرات على شكل غلايكوجين ويقود تحلله في النهاية إلى إطلاق الجلوكوز من خلايا الكبد، وذلك أمر ضروري لتنظيم تركيز السكر في الدم. الناتج الأولي لتحلل الغلايكوجين هو غلوكوز 1-فوسفات ويتم تحويله إلى غلوكوز 6-فوسفات ثم يقوم إنزيم في الشبكة الملساء لخلية الكبد بإزالة الفوسفات من الغلوكوز ليتمكن بعدها من مغادرة الخلية.[25][30]
يمكن لإنزيمات الشبكة الملساء المساعدة في إزالة سُمّية الأدوية. يتم في العادة أثناء إزالة السمية إضافة مجموعة هيدروكسيل للدواء تجعله أكثر قابلية للذوبان ومنه إزالته من الجسد. أحد التفاعلات المدروسة بشكل كبير يتم بواسطة عائلة سيتوكروم بي450 من الإنزيمات، والتي تحفز هيدركسلة الأدوية غير القابلة للذوبان في الماء أو المستقلبات التي كانت -من دون ذلك- لتتراكم إلى مستويات سُمّية في غشاء الخلية. للشبكة الملساء الخاصة بالخلايا العضلية وظيفة اختصاصية أخرى، حيث يقوم غشاء الشبكة الملساء فيها بضخ أيونات الكالسيوم من العصارة الخلوية إلى الفراغ الصهريجي. حين تصبح الخلية العضلية منبه بواسطة تدفع عصبي، يرجع الكالسيوم عبر غشاء الشبكة إلى العصارة الخلوية ويولد تقلص الخلية العضلية.[25][30]
وظائف الشبكة الإندوبلازمية الخشنة
تصدِّر العديد من أنواع الخلايا البروتينات المخلقة بواسطة الريبوسومات المرتبطة بالشبكة الإندوبلازمية الخشنة (RER)، حيث تُجمِّع الريبوسومات الأحماض الأمينية إلى وحدات بروتينية ويتم نقلها إلى الشبكة الخشنة لإجراء تعديلات إضافية لها. يمكن لهذه البروتينات أن تكون إما: بروتينات عبر غشائية يتم دمجها في غشاء الشبكة الإندوبلازمية، وإما بروتينات قابلة للذوبان في الماء يمكنها العبور عبر الغشاء إلى اللمعة. تُطوى البروتينات التي تصل إلى داخل الشبكة الإندوبلازمية إلى بنياتها الثالثية الصحيحة، وتضاف إليها مركبات كيميائية مثل الكربوهيدرات أو السكريات، وبعدها إما أن تنقل الشبكة الإندوبلازمية البروتينات الكاملة -والتي تسمى بروتينات مفرزة- إلى مناطق مختلفة من الخلية تحتاج إليها، وإما أن تُرسل إلى جهاز غولجي للمزيد من المعالجة والتعديلات.[25][30]
بعد تشكُّل البروتينات المفرزة، يقوم غشاء الشبكة الإندوبلازمية بفصلها عن البروتينات التي ستبقى في العصارة الخلوية. تغادر البروتينات المفرزة من الشبكة الإندوبلازمية ملفوفة في أغشية حويصلات تتبرعم مثل الفقاعات من الشبكة الإندوبلازمية الانتقالية، تسمى هذه الحويصلات التي تنتقل إلى جزء آخر من الخلية بالحويصلات الناقلة.[25][30] آلية أخرى لنقل الليبيدات والبروتينات خارج الشبكة الإندوبلازمية تحدث عبر البروتينات الناقلة للبيد في مناطق تسمى مواقع اتصال غشائية تصبح فيها الشبكة الإندوبلازمية مرتبطة على مسافة قريبة وثابتة بأغشية عضيات أخرى مثل الغشاء الخلوي، جهاز غولجي واليحلولات.[31]
إضافة إلى تكوين البروتينات المفرزة، تكوِّن الشبكة الإندوبلازمية أغشية تنمو فيها بإضافة البروتينات والليبيدات الفوسفورية. بينما تنمو عديدات الببتيد التي يُفترض أن تصبح بروتينات غشائية في الريبوسوم، يتم إدراجها داخل غشاء الشبكة الإندوبلازمية وإبقاؤها هنالك بواسطة أجزائها الكارهة للماء. تُنتِج الشبكة الخشنة كذلك ليبيداتها الفوسفورية الخاصة بها، وتقوم الإنزيمات المدمجة في غشاء الشبكة الإندوبلازمية بتجميع الليبيدات الفوسفورية. يتمدد غشاء الشبكة الإندوبلازمية ويمكن أن يتم نقله بواسطة الحويصلات الناقلة إلى مكونات أخرى للنظام الغشائي الداخلي.[25][30]
جهاز غولجي

يتكون جهاز غولجي (ويعرف كذلك بجسم غولجي ومركب غولجي) من جيوب منفصلة تسمى صهاريج، وشكلها مماثل لحزمة من الفطائر المحلاة. ويختلف عدد هذه الحزمات على حسب اختصاص الخلية الوظيفي. يُستخدم جهاز غولجي من قبل الخلية لإجراء المزيد من التعديلات على البروتينات المخلقة حديثا، يسمى الجزء الذي يستقبل الحويصلات من جهاز غولجي بالوجه مقرون ويكون عادة بجوار الشبكة الإندوبلازمية، وتسمى النهاية المعاكسة للوجه مقرون من جهاز غولجي بالوجه مفروق ومنه تغادر المركبات الكاملة، ويقابل الوجه مفروق الغشاء الخلوي عادة، وإليه تُرسل معظم المواد التي يعدّلها جهاز غولجي.[32]
تخضع البروتينات التي تُرسلها الشبكة الإندوبلازمية في حويصلاتٍ إلى المزيد من التعديلات في جهاز غولجي وبعدها يتم التحضير لإفرازها خارج الخلية أو لنقلها إلى أماكن أخرى منها. يمكن أن تحدث العديد من الأمور للبروتينات أثناء رحلتها في فراغ جهاز غولجي المغطى بالإنزيمات، حيث يزيل الجهاز موحودات سكر بديلة منتجًا مجموعةً كبيرةً متنوعة من قليلات السكاريد. علاوة على تعديل البروتينات، يركب جهاز غولجي جزيئات ضخمة كذلك ففي الخلايا النباتية ينتج جهاز غولجي بكتين والعديد من عديدات السكاريد الأخرى التي تحتاج إليها.[33]
عند انتهاء عملية التعديلات الإضافية، يقوم جهاز غولجي بفرز النواتج التي عالجها وإرسالها إلى أجزاء مختلفة من الخلية بعد إضافة لصاقات تعرُّف جزيئية أو واسمات بروتينية لها لمساعدة العضيات المستقبلة لها في التعرف عليها. بعد تنظيم كل شيء يُرسل جهاز غولجي نواتجه عبر حويصلات متبرعمة من الوجه المفروق.[34]
الفجوات العصارية
| علم الأحياء الخلوي | |
|---|---|
| الخلية الحيوانية | |
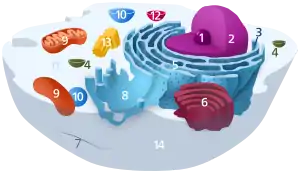 مكونات الخلية الحيوانية النموذجية:
| |
الفجوات العصارية مثل الحويصلات هي أكياس غشائية (محاطة بغشاء) داخل الخلية، وهي أكبر حجما من الحويصلات وذات وظائف متخصصة متعددة. وظائف الفجوات العصارية تختلف بين الخلايا النباتية والحيوانية.
في الخلايا النباتية، تشغل الفجوات العصارية حجما يتراوح بين 30% إلى 90% من الحجم الكلي للخلية.[35] معظم الخلايا النباتية البالغة تحتوي على فجوة عصارية مركزية واحدة يحيط بها غشاء يسمى غشاء الفجوة (tonoplast). تعمل الفجوات في الخلايا النباتية كأحيازٍ لتخزين المغذيات وفضلات الخلية، ويسمى المحلول الذي تُخزَّن فيه هذه الجزيئات بالعصارة الخلوية. تتواجد الخضابات التي تُلون الخلية في بعض الأحيان بالعصارة الخلوية. يمكن للفجوات زيادة حجم الخلية -التي تتمدد بإضافة الماء- كما أنها تتحكم في ضغط الامتلاء (الضغط الإسموزي الذي يحول دون انحناء جدار الخلية للداخل). مثل اليحلولات في الخلايا الحيوانية، لدى الفجوات العصارية أس هيدروجيني حمضي وتحتوي على إنزيمات حلمأة. يمكِّن الأس الهيدروجيني الفجوات من القيام بإجراءات استتبابية في الخلية، على سبيل المثال حين ينخفظ الأس الهيدروجيني في محيط الخلايا يمكن تحويل أيونات +H المتدفقة إلى العصارة الخلوية نحو فجوة عصارية للحفاظ على ثابت الأس الهيدروجيني للعصارة الخلوية.[36]
في الخلايا الحيوانية، تساهم الفجوات في عمليتي الإيماس والالتقام الخلوي. يشير الالتقام الخلوي إلى عملية إدخال المواد داخل الخلية ويشير الإيماس إلى عملية إخراج المواد من الخلية إلى الوسط خارج الخلوي. تحاط المواد المُدخلة إلى داخل الخلية بالغشاء الخلوي وبعدها تنقل إلى فجوة عصارية، يوجد نوعان من الالتقام الخلوي: البلعمة (ابتلاع الخلايا) والاحتساء (شرب الخلايا). تقوم الخلايا في البلعمة بإحاطة جزيئات كبيرة مثل البكتيريا. الاحتساء عملية مماثلة باستثناء أن الجزيئات التي يتم ابتلاعها تكون في حالة سائلة.[37]
الحويصلات
الحويصلات عبارة عن وحدات نقل صغيرة مكونة من أغشية مغلقة يمكنها نقل الجزيئات بين مختلف الأحياز الخلوية. تَنقل معظم الحويصلات الأغشيةَ المجمَّعَةَ في الشبكة الإندوبلازمية إلى جهاز غولجي، ثم من جهاز غولجي إلى مواقع مختلفة.[38]
تتواجد أنواع عديدة من الحويصلات ويحتوي كل نوع منها على بروتينات مختلفة في التكوين. تتشكل معظم هذه الحويصلات من مناطق محددة من الأغشية. حين تتبرعم الحويصلة من غشاء ما فإنها تحتوي بروتينات محددة في سطح عصارتها الخلوية. يحتوي كل غشاء تسافر إليه الحويصلة واسما في سطح عصارته الخلوية، يتطابق هذا الواسم مع البروتينات التي في الحويصلة المسافرة إلى هذا الغشاء. وعندما تجد الحويصلة الغشاء المتطابق تندمج معه.[39]
توجد ثلاث أنواع شائعة من الحويصلات هي: الحويصلات المغلفة بالكلاثرين، الحويصلات المغلفة بـ COPI والحويصلات المغلفة بـ COPII ويؤدي كل منها وظائف مختلفة في الخلية، مثلا: تنقل الحويصلات المغلفة بالكلاثرين المواد بين جهاز غولجي والغشاء الخلوي في حين أن الحويصلات المغلفة بـ COPI وCOPII تستخدمان بشكل متواتر في نقل المواد بين الشبكة الإندوبلازمية وجهاز غولجي.[39]
الأجسام الحالة
اليحلولات أو الجسيمات الحالة هي عضيات تحتوي على إنزيمات حلمأة تُستخدم في الهضم داخل الخلوي. الوظيفة الأساسية لليحلول هي معالجة الجزيئات التي تُدخلها الخلية إلى وسطها وإعادة تدوير أجزاء الخلية المتضررة والمهترئة. الإنزيمات المتواجدة داخل اليحلولات هي هيدرولازات حمضية تتطلب أوساطا حمضية لأدائها المثالي. توفر اليحلولات أوساطا مثل تلك بالحفاظ على أس هيدروجيني يساوي 5.0 داخل العضية.[40] إذا حدث وتمزق اليحلول فإن الإنزيمات المحررة لا تكون نشيطة جدا لأن الأس الهيدروجيني للعصارة الخلوية متعادل. لكن إذا تمزقت عدة يحلولات وتسربت إنزيماتها فيمكن أن يسبب ذلك انهضاما ذاتيا وتتدمر الخلية.
تقوم اليحلولات بالهضم داخل الخلوي - بواسطة عملية تسمى البعلمة- وذلك عبر الاندماج مع فجوة عصارية وتحرير إنزيماتها داخل الفجوة. بفضل هذه العملية تمر السكريات والأحماض الأمينية وموحدات أخرى إلى العصارة الخلوية وتصبح مغذيات ومواد أولية للخلية، تستخدم اليحلولات إنزيمات الحلمأة خاصتها لإعادة تدوير عضياتها القديمة والمهترئة عبر عملية تسمى الالتهام الذاتي، حيث يحيط اليحلول بعضية مهترئة ويستخدم إنزيماته لتفكيك المادة المهضومة، ينتج عن ذلك موحودات عضوية تعود إلى العصارة الخلوية لإعادة استخدامها. المهمة الأخيرة لليحلول هي هضم الخلية نفسها عبر عملية تسمى التحلل الذاتي.[41]
الجسم المدبب

الجسم المدبب أو الجسم الطرفي هو أحد مكونات النظام الغشائي الداخلي ويتواجد في الفطريات فقط، وله علاقة بنمو الخيوط الفطرية، وهو جسيم مرتبط بمرحلة الظلام ويتكون من تكدس فجوات غشائية تحتوي على مكونات جدار خلوي، ويعمل كنقطة تجميع وتحرير لمثل هذه المكونات التي تتوسط جهاز غولجي والغشاء الخلوي. الجسيم القمي هو عضية حركية ويولد خيوطا فطريا مع تحركه إلى الأمام.[10]
الغشاء الخلوي
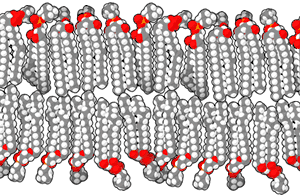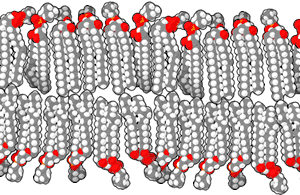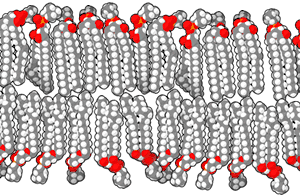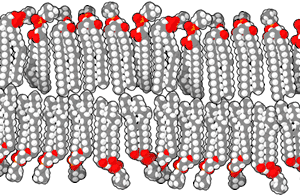
الغشاء الخلوي أو الغشاء البلازمي هو غشاء ذو طبقة مزدوجة من الليبيدات الفوسفورية يفصل الخلية عن محيطها الخارجي وينظم نقل الجزيئات والإشارات إلى داخل وخارج الخلية. توجد في الغشاء بروتينات مدمجة هي التي تؤدي الوظائف التي يقوم بها الغشاء. الغشاء البلازمي ليس بنية ثابتة أو صلبة، إذ يمكن للجزيئات التي تكونه التحرك جانبيا، بسبب هذه الحركة ومكوناته المتعددة الأخرى يُشار إلى الغشاء البلازمي بأنه فسيفساء مائعة. يمكن للجزيئات الأصغر مثل ثنائي أكسيد الكربون، الماء والأكسجين المرور عبر الغشاء البلازمي بحرية بواسطة الانتشار أو التناضح، أما الجزيئات الكبيرة التي تحتاجها الخلية فيتم نقلها بمساعدة البروتينات عبر النقل النشط.[42]
يقوم الغشاء البلازمي الخاص بالخلية بعدة وظائف منها: نقل المغذيات إلى الخلية، السماح بمغادرة الفضلات، منع المواد الدخيلة من دخول الخلية، تفادي مغادرة المواد التي تحتاجها الخلية، الحفاظ على الأس الهيدروجيني الخاص بالعصارة الخلوية ومنع الضغط الإسموزي للعصارة الخلوية. البروتينات الناقلة التي تسمح لبعض المواد بالعبور من خلالها ولا تسمح للبعض الآخر تُستخدم لهذه الوظائف، تستخدم هذه البروتينات حلمأة الـATP لضخ المواد ضد تدرجات تركيزها.[42]
فضلا عن هذه الوظائف العامة، لدى الغشاء البلازمي وظائف محددة في الكائنات متعددة الخلايا، حيث تساعد البروتينات السكرية المتواجدة في الغشاء الخليَةَ في التعرف على خلايا أخرى في سبيل تبادل المستقلبات وتشكيل الأنسجة. تسمح بروتينات أخرى في الغشاء البلازمي بالارتباط بالهيكل الخلوي والنسيج البين خارج الخلوي، وهي وظيفة تحافظ على شكل الخلية وتثبت مواقع البروتينات الغشائية. الإنزيمات التي تحفز التفاعلات توجد كذلك في الغشاء البلازمي. لدى البروتينات المستقبلة في الغشاء شكل يطابق الجزيء الكيميائي الرسول، وهذا ينتج عنه استجابات خلوية متعددة.[43]
التطور
يرتبطُ أصل النظام الغشائي الداخلي مع أصل حقيقيات النوى نفسها وأصل حقيقات النوى مع أصل الميتوكندريا المُتعايشة داخليًا. طُرحت العديد من النماذج لمحاولة شرح أصل النظام الغشائي الداخلي.[44] يُشير التصور الأحدث إلى أنَّ النظام الغشائي الداخلي قد تطورَ من حويصلات الغشاء الخارجي التي تُفرزها الميتوكندريا المُتعايشة داخليًا.[45] حاليًا، يُعتبر النموذج المبني على حويصلات الغشاء الخارجي (OMV-based model) والذي يُوضح أصل النظام الغشائي الداخلي، النموذجَ الذي يتطلب أقلَ قدرٍ من الاكتشافات الجديدة في أصل حقيقيات النوى، حيثُ أنهُ يشرح العديد من روابط الميتوكندريا مع الأجزاء الأخرى للخلية.[46]
المراجع
- "Al-Qamoos القاموس - Biology - Endomembrane system"، www.alqamoos.org، مؤرشف من الأصل في 6 مارس 2019، اطلع عليه بتاريخ 2 مارس 2019.
- "ترجمة و معنى endomembrane system بالعربي في قاموس المعاني. قاموس عربي انجليزي مصطلحات صفحة 1"، www.almaany.com، مؤرشف من الأصل في 6 مارس 2019، اطلع عليه بتاريخ 2 مارس 2019.
- Smith, A. L. (1997)، Oxford dictionary of biochemistry and molecular biology، Oxford [Oxfordshire]: Oxford University Press، ص. 206، ISBN 0-19-854768-4.
- Davidson, Michael (2005)، "The Nuclear Envelope"، Molecular Expressions، Florida State University، مؤرشف من الأصل في 16 مايو 2019، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Davidson, Michael (2005)، "The Endoplasmic Reticulum"، Molecular Expressions، Florida State University، مؤرشف من الأصل في 15 مايو 2019، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Graham, Todd R. (2000)، Eurekah Bioscience Collection Cell Biology، University of New South Wales and Landes Bioscience، ISBN 0-7334-2108-3، مؤرشف من الأصل في 14 أبريل 2020.
- Lodish, Harvey؛ وآخرون (2000)، "Section 5.4 Organelles of the Eukaryotic Cell"، Molecular Cell Biology، W. H. Freeman and Company، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Cooper, Geoffrey (2000)، "The Mechanism of Vesicular Transport"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Davidson, Michael (2005)، "Plasma Membrane"، Molecular Expressions، Florida State University، مؤرشف من الأصل في 15 مايو 2019، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Steinberg, G. (2007)، "Hyphal Growth: a Tale of Motors, Lipids, and the Spitzenkörper"، Eukaryotic Cell، 6 (3): 351–360، doi:10.1128/EC.00381-06، PMC 1828937، PMID 17259546، مؤرشف من الأصل في 14 أكتوبر 2011.
- "Prokaryotic photosynthesis and phototrophy illuminated"، Trends Microbiol.، 14 (11): 488–96، 2006، doi:10.1016/j.tim.2006.09.001، PMID 16997562.
- "Lamellar Organization of Pigments in Chlorosomes, the Light Harvesting Complexes of Green Photosynthetic Bacteria"، Biophys. J.، 87 (2): 1165–72، أغسطس 2004، doi:10.1529/biophysj.104.040956، PMC 1304455، PMID 15298919، مؤرشف من الأصل في 10 مايو 2020.
- Campbell, Neil A.؛ Reece, Jane B. (2002)، Biology (ط. 6th)، Benjamin Cummings، ISBN 0-8053-6624-5، مؤرشف من الأصل في 25 أكتوبر 2019.
- "Phospholipid synthesis and lipid composition of subcellular membranes in the unicellular eukaryote Saccharomyces cerevisiae"، J. Bacteriol.، 173 (6): 2026–34، مارس 1991، PMC 207737، PMID 2002005، مؤرشف من الأصل في 16 مارس 2020.
- "Synthesis, storage and degradation of neutral lipids in yeast"، Biochim. Biophys. Acta، 1771 (3): 299–309، مارس 2007، doi:10.1016/j.bbalip.2006.07.001، PMID 16916618.
- Futerman AH (ديسمبر 2006)، "Intracellular trafficking of sphingolipids: relationship to biosynthesis"، Biochim. Biophys. Acta، 1758 (12): 1885–92، doi:10.1016/j.bbamem.2006.08.004، PMID 16996025.
- "Biochemistry of mammalian peroxisomes revisited"، Annu. Rev. Biochem.، 75: 295–332، 2006، doi:10.1146/annurev.biochem.74.082803.133329، PMID 16756494.
- Voelker DR (1 ديسمبر 1991)، "Organelle biogenesis and intracellular lipid transport in eukaryotes"، Microbiol. Rev.، 55 (4): 543–60، PMC 372837، PMID 1779926، مؤرشف من الأصل في 16 مارس 2020.
- Voelker DR (يوليو 2005)، "Bridging gaps in phospholipid transport"، Trends Biochem. Sci.، 30 (7): 396–404، doi:10.1016/j.tibs.2005.05.008، PMID 15951180.
- Morré DJ, Mollenhauer HH. (1974). The endomembrane concept: a functional integration of endoplasmic reticulum and Golgi apparatus. In Dynamic Aspects of Plant infrastructure (ed. A. W. Robards), pp. 84–137. London, New York, etc.: McGraw-Hill.
- Morre, D.J. (1975)، "Membrane Biogenesis"، Annual Review of Plant Physiology، 26 (1): 441–481، doi:10.1146/annurev.pp.26.060175.002301.
- Childs, Gwen V. (2003)، "Nuclear Envelope"، UTMB، مؤرشف من الأصل في 20 يونيو 2006، اطلع عليه بتاريخ 28 سبتمبر 2008.
- Cooper, Geoffrey (2000)، "The Nuclear Envelope and Traffic between the Nucleus and Cytoplasm"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 09 مايو 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Alberts, Walter؛ وآخرون (2002)، "Nuclear Pore Complexes Perforate the Nuclear Envelope"، Molecular Biology of the Cell 4th edition، Garland Science، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Cooper, Geoffrey (2000)، "The Endoplasmic Reticulum"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Bertolotti, Anne؛ Zhang, Yuhong؛ Hendershot, Linda M.؛ Harding, Heather P.؛ Ron, David (2000)، "Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response"، Nature Cell Biology، 2 (6): 326–333، doi:10.1038/35014014، PMID 10854322، مؤرشف من الأصل في 14 يونيو 2016، اطلع عليه بتاريخ 03 أكتوبر 2008.
- Protein translocation across the ER membrane نسخة محفوظة 11 أغسطس 2012 على موقع واي باك مشين.
- Entry into the Endoplasmic Reticulum: Protein Translocation, Folding and Quality Control نسخة محفوظة 17 ديسمبر 2019 على موقع واي باك مشين.
- Biology، McGraw Hill education، 2011، ص. 89.
- Alberts, Walter؛ وآخرون (2002)، "Membrane-bound Ribosomes Define the Rough ER"، Molecular Biology of the Cell 4th edition، Garland Science، مؤرشف من الأصل في 10 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- "Inter-organelle membrane contact sites: through a glass, darkly"، Curr. Opin. Cell Biol.، 18 (4): 371–8، أغسطس 2006، doi:10.1016/j.ceb.2006.06.011، PMID 16806880.
- Rothman, J. (1981)، "The golgi apparatus: two organelles in tandem"، Science، 213 (4513): 1212–1219، doi:10.1126/science.7268428، PMID 7268428، مؤرشف من الأصل في 09 مايو 2020، اطلع عليه بتاريخ 04 أكتوبر 2008.
- Alberts, Walter؛ وآخرون (2002)، "Transport from the ER through the Golgi Apparatus"، Molecular Biology of the Cell 4th edition، Garland Science، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Cooper, Geoffrey (2000)، "The Golgi Apparatus"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Alberts, Walter؛ وآخرون (2002)، "Plant and Fungal Vacuoles Are Remarkably Versatile Lysosomes"، Molecular Biology of the Cell 4th edition، Garland Science، مؤرشف من الأصل في 10 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Lodish, Harvey؛ وآخرون (2000)، "Plant Vacuoles Store Small Molecules and Enable the Cell to Elongate Rapidly"، Molecular Cell Biology، W. H. Freeman and Company، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Cooper, Geoffrey (2000)، "Endocytosis"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Lodish, Harvey؛ وآخرون (2000)، "Section 17.10 Molecular Mechanisms of Vesicular Traffic"، Molecular Cell Biology، W. H. Freeman and Company، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Alberts, Walter؛ وآخرون (2002)، "The Molecular Mechanisms of Membrane Transport and the Maintenance of Compartmental Diversity"، Molecular Biology of the Cell 4th edition، Garland Science، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Alberts, Walter؛ وآخرون (2002)، "Transport from the Trans Golgi Network to Lysosomes"، Molecular Biology of the Cell 4th edition، Garland Science، مؤرشف من الأصل في 10 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Cooper, Geoffrey (2000)، "Lysosomes"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Cooper, Geoffrey (2000)، "Structure of the Plasma Membrane"، The Cell: A Molecular Approach، Sinauer Associates, Inc، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Lodish, Harvey؛ وآخرون (2000)، "Section 5.3. Biomembranes: Structural Organization and Basic Functions"، Molecular Cell Biology، W. H. Freeman and Company، مؤرشف من الأصل في 14 أبريل 2020، اطلع عليه بتاريخ 09 ديسمبر 2008.
- Martin WF وآخرون (6 مايو 2014)، "Endosymbiotic theories for eukaryote origin"، Philos. Trans. R. Soc. B، 370: 20140330–18، doi:10.1098/rstb.2014.0330، PMC 4571569، PMID 26323761، مؤرشف من الأصل في 8 سبتمبر 2018.
- Gould SB وآخرون (31 مارس 2016)، "Bacterial Vesicle Secretion and the Evolutionary Origin of the Eukaryotic Endomembrane System"، Trends Microbiol.، 24: 525–34، doi:10.1016/j.tim.2016.03.005، PMID 27040918، مؤرشف من الأصل في 17 ديسمبر 2019.
- Murley, A.؛ Nunnari, J. (03 مارس 2016)، "The emerging network of mitochondria-organelle contacts"، Mol. Cell، 61: 648–653، doi:10.1016/j.molcel.2016.01.031، PMID 26942669، مؤرشف من الأصل في 17 ديسمبر 2019.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء