بروتين الفصل
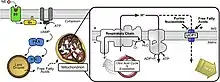
بروتين الفصل ( UCP ) هو من بروتينات الغشاء الداخلي للميتوكوندريا وهو عبارة عن قناة أو بروتون منظم. يستطيع بروتين الفصل أيضاً تفريق ممال البروتونات المُولد بضخ البروتونات بطاقة NADH من المطرس الميتوكوندري إلى الفضاء الميتوكوندري داخل الغشائي لا تُستخدم الطاقة المفقودة في تفريق ممال البروتونات عبر بروتينات الفصل بالعمل الكيميائي الحيوي. بدلاً من ذلك ، يتم توليد الحرارة. هذا ما يربط UCP بالتوليد الحراري. ومع ذلك ، لا ترتبط جميع أنواع UCPs بالحرارة. على الرغم من أن UCP2 و UCP3 يرتبطان ارتباطًا وثيقًا بـ UCP1 ، فإن UCP2 و UCP3 لا يؤثران على قدرات التنظيم الحراري للفقاريات. [1] تتموضع بروتينات الفصل في نفس غشاء ATP synthase وهو قناة بروتون أيضاً.يعمل البروتينان بالتوازي بحيث يولد أحدهما الحرارة ويولد الآخر ATP من ADP والفوسفات غير العضوية، وهي المرحلة الأخيرة من الفسفرة التأكسدية . [2] يرتبط التنفس الميتوكوندري بتركيب ATP (فسفرة ADP) لكنه تنظمه بروتينات الفصلUCP. [3]

تؤدي بروتينات الفصل دوراً في الفيزيولوجيا الطبيعية كما في التعرض للبرد أو في السبات لأن الطاقة تُستخدم في توليد الحرارة (انظر في توليد الحرارة) بدلاً من إنتاج ATP. تستخدم بعض أنواع النبات الحرارة المتولدة عن بروتينات الفصل لأغراض خاصة. يبقي ملفوف الظريبان مثلاً حرارة أشواكه أعلى بعشرين درجة مئوية من البيئة المحيطة لينشر الرائحة ويجذب الحشرات التي تلقح الأزهار. [4] تؤدي مواد أخرى مثل 2,4-dinitrophenol وcarbonyl cyanide m-chlorophenyl hydrazone الوظيفة ذاتها. حمض الساليسيليك عامل فصل أيضاً (في النباتات بشكل رئيس) كما أنه يخفض إنتاج ATP ويرفع حرارة الجسم إذا أُخذ بزيادة كبيرة. [5] تزداد بروتينات الفصل بالهرمون الدرقي والنورابينفرين والإبنفرين واللبتين. [6]
التاريخ
لاحظ العلماء نشاطا حراريا في الأنسجة الدهنية البنية ، مما أدى في النهاية إلى اكتشاف UCP 1 ، المعروف في البداية باسم «بروتين Uncoupling». [3] أظهر النسيج البني نسباً مرتفعة من التنفس الميتوكوندري وتنفس آخر غير مرتبط بتركيب ATP ، ما رمز إلى نشاط حراري قوي. كان UCP1 البروتين المكتشف المسؤول عن تفعيل سبيل البروتين الغير مرتبط بفسفرة ADP (الذي يحدث عادة عبر ATP synthase).
في الثدييات
هناك خمسة نظائر UCP معروفة عند الثدييات. في حين يمثل كل واحد منها وظيفة مميزة، فإن بعض الوظائف يؤديها أكثر من مماثل منها. المماثلات كالتالي:
- UCP1 ، المعروف أيضًا باسم thermogenin
- UCP2 ، المعروف أيضا باسم SLC25A8
- UCP3 ، والمعروف أيضًا باسم SLC25A9
- UCP 4 ، والمعروف أيضًا باسم SLC25A27
- UCP 5 ، والمعروف أيضًا باسم SLC25A14
الحفاظ على درجة حرارة الجسم
اكتشف أول بروتين فصل UCP1 في النسيج الدهني البني في حيوانات السبات الشتوي والقوارض الصغيرة ما يؤمن حرارة غير ارتعاشية لهذه الحيوانات. [3] هذه النسج الدهنية البنية أساسية للحفاظ على حرارة جسد القوارض الصغيرة وتظهر الدراسات على الفئران المعدلة بأن هذه النُسج لا تعمل بشكل صحيح بدون بروتينات فصل فعالة. أظهرت هذه الدراسات في الحقيقة أن التأقلم مع البرد غير ممكن لدى هذه الفئران المعدلة ما يشير إلى أن UCP1 عنصر أساسي لإنتاج الحرارة في هذه النسج الدهنية البنية. [7] [8]
في مناطق أخرى من الجسم، من المعروف تأثير نشاط بروتينات الفصل على الحرارة في البيئات الصغروية. [9] [10] يُعتقد بتأثير ذلك على نشاطات البروتينات الأخرى في هذه المناطق. لكن ما زال هناك حاجة لمزيد من العمل لتحديد النتائج الحقيقية لممالات الحرارة الناجمة عن الفصل في الخلايا.
الدور في تراكيز ATP
يختلف تأثير UCP2 وUCP3 على تراكيز ATP اعتماداً على نمط الخلية. [9] تظهر خلايا بيتا البنكرياسية على سبيل المثال نقصاً في ATP وزيادة في UCP2. يرتبط ذلك بالتنكس الخلوي وانخفاض إفراز الأنسولين والسكري من النمط الثاني. [11] على العكس، يحفز UCP2في الحصين وUCP3 في الخلايا العضلية إنتاج الميتوكوندريا. [12] زيادة عدد الميتوكوندريا تزيد تركيز ADP وATP ما يؤدي فعلاً إلى زيادة صافية في تركيز ATP عندما ترتبط بروتينات الفصل هذه( تتثبط آلية السماح بتسريب البروتونات).
صياغة تركيز أنواغ الأوكسجين التفاعلية
القائمة الكاملة لوظائف UCP 2 و UCP 3 غير معروفة. [13] لكن تظهر الدراسات أن هذه البروتينات تتدخل في حلقة تلقيم راجع سلبي تحد من تركيز أنواع الأكسجين التفاعلية أنواع الأكسجين التفاعلية (ROS). [14] يشير الإجماع العلمي الحالي إلى أن UCP 2 و UCP 3 يقومان بنقل البروتونات فقط عند وجود أنواع التنشيط . [15] من بين هذه المنشطات الأحماض الدهنية ، ROS ، وبعض نواتج ROS المرافقة التفاعلية أيضاً. لبذلك تسبب النسب المرتفعة من ROS تزايداً في نشاط UCP2 وUCP3. يزيد ذلك بدوره تسرب البروتونات من الميتوكوندريا ما يخفض من قوة جذب البروتون عبر غشاء الميتوكوندريا ويفعل سلسلة نقل الإلكترون. تقليل القوة الدافعة للبروتون بهذه العملية تسبب حلقة تخفيض راجع سلبي تقلل من إنتاج ROS. على وجه الخصوص ، يقلل UCP2 من قدرة الغشاء الميتوكوندريا عبر الغشاء ، وبالتالي يقلل من إنتاج ROS. وبالتالي ، قد تزيد الخلايا السرطانية من إنتاج UCP2 في الميتوكوندريا. [16] يدعم هذه النظرية دراسات مستقلة تظهر زيادة إنتاج ROS في كل من الفئران المعدلة بحيث حذف منها UCP 2 و UCP 3 .
ذه العملية مهمة لصحة الإنسان للاعتقاد بمساهمة تراكيز ROS المرتفعة بتطور الأمراض التنكسية. [15]
الوظائف في العصبونات
عن طريق الكشف عن مرنا المرتبطة ،ظهر انتشار UCP 2 ، UCP 4 ، و UCP 5 في الخلايا العصبية في جميع أنحاء الجهاز العصبي المركزي البشري. [17] تلعب هذه البروتينات أدوارًا رئيسية في وظيفة الخلايا العصبية. [9] في حين أن العديد من نتائج الدراسة لا تزال موضع جدل ، الا أن بعضها مقبول على نطاق واسع.
على سبيل المثال ، يغير UCP تركيز الكالسيوم الحر في العصبون. [9] الميتوكوندريا هي مخزن رئيس للكالسيوم في العصبونات، وتتزايد السعة التخزينية مع وجود كمون عبر أغشية الميتوكوندريا. [18] لذلك عندما تقلل بروتينات الفصل الكمون عبر هذه الأغشية، تتحرر أيونات الكالسيوم إلى البيئة المحيطة في العصبون. بسبب ارتفاع تركيز الميتوكوندريا قرب نهايات المحاور، يدل ذلك على دور UCP في تنظيم تراكيز الكالسيوم في هذه المنطقة. باعتبار دور أيونات الكالسيوم الكبير في النقل العصبي، يتوقع العلماء أن هذه البروتينات UCP تؤثر في النقل العصبي مباشرة.
كما ذُكر في الأعلى، تشهد العصبونات في الحصين ازدياداً في تراكيز ATP بوجود بروتينات الفصل هذه. [9] [12] يقود هذا العلماء للافتراض بأن UCP يحسّن الليونة والنقل المشبكيين.
المراجع
- Gaudry, Michael J.؛ Jastroch, Martin (مارس 2019)، "Molecular evolution of uncoupling proteins and implications for brain function"، Neuroscience Letters، 696: 140–145، doi:10.1016/j.neulet.2018.12.027، ISSN 0304-3940، مؤرشف من الأصل في 18 مايو 2020.
- "Uncoupling proteins: current status and therapeutic prospects"، EMBO Reports، 6 (10): 917–21، أكتوبر 2005، doi:10.1038/sj.embor.7400532، PMID 16179945.
- "The biology of mitochondrial uncoupling proteins"، Diabetes، 53 (suppl 1): S130-5، فبراير 2004، doi:10.2337/diabetes.53.2007.S130، PMID 14749278.
- Garrett, Reginald H.؛ Grisham, Charles M. (2013)، Biochemistry (ط. Fifth Edition, International)، China: Mary Finch، ص. 668، ISBN 978-1-133-10879-5.
- "California Poison Control System: Salicylates"، مؤرشف من الأصل في 2 أغسطس 2014، اطلع عليه بتاريخ 17 سبتمبر 2020.
- "Uncoupling protein-3 is a mediator of thermogenesis regulated by thyroid hormone, beta3-adrenergic agonists, and leptin"، The Journal of Biological Chemistry، 272 (39): 24129–32، سبتمبر 1997، doi:10.1074/jbc.272.39.24129، PMID 9305858.
- "Mitochondrial uncoupling proteins in human physiology and disease"، Minerva Medica، 93 (1): 41–57، فبراير 2002، PMID 11850613.
- "UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality"، Cell Metabolism، 9 (2): 203–9، فبراير 2009، doi:10.1016/j.cmet.2008.12.014، PMID 19187776.
- "Mitochondrial uncoupling proteins in the CNS: in support of function and survival"، Nature Reviews. Neuroscience، 6 (11): 829–40، نوفمبر 2005، doi:10.1038/nrn1767، PMID 16224498.
- "Brain uncoupling protein 2: uncoupled neuronal mitochondria predict thermal synapses in homeostatic centers"، The Journal of Neuroscience، 19 (23): 10417–27، ديسمبر 1999، PMID 10575039.
- "Uncoupling protein-2 negatively regulates insulin secretion and is a major link between obesity, beta cell dysfunction, and type 2 diabetes"، Cell، 105 (6): 745–55، يونيو 2001، doi:10.1016/s0092-8674(01)00378-6، PMID 11440717.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Uncoupling protein 2 prevents neuronal death including that occurring during seizures: a mechanism for preconditioning"، Endocrinology، 144 (11): 5014–21، نوفمبر 2003، doi:10.1210/en.2003-0667، PMID 12960023.
- "Mitochondrial proton and electron leaks"، Essays in Biochemistry، 47: 53–67، 14 يونيو 2010، doi:10.1042/bse0470053، PMID 20533900.
- "Uncoupling proteins and the control of mitochondrial reactive oxygen species production"، Free Radical Biology & Medicine، 51 (6): 1106–15، سبتمبر 2011، doi:10.1016/j.freeradbiomed.2011.06.022، PMID 21762777.
- "Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3"، Cell Metabolism، 2 (2): 85–93، أغسطس 2005، doi:10.1016/j.cmet.2005.06.002، PMID 16098826.
- Sreedhar, Annapoorna؛ Zhao, Yunfeng (مايو 2017)، "Uncoupling protein 2 and metabolic diseases"، Mitochondrion، 34: 135–140، doi:10.1016/j.mito.2017.03.005، ISSN 1567-7249، مؤرشف من الأصل في 18 مايو 2020.
- "Uncoupling protein 2 in the brain: distribution and function"، Biochemical Society Transactions، 29 (Pt 6): 812–7، نوفمبر 2001، doi:10.1042/bst0290812، PMID 11709080.
- "Mitochondrial membrane potential and neuronal glutamate excitotoxicity: mortality and millivolts"، Trends in Neurosciences، 23 (4): 166–74، أبريل 2000، doi:10.1016/s0166-2236(99)01534-9، PMID 10717676.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية