بروتين دمج الغشاء
بروتين دمج الغشاء أو بروتين التحام الغشاء (بالإنجليزية: Membrane fusion protein) هي بروتينات تسبب اندماج الأغشية الحيوية [الإنجليزية]. اندماج الأغشية أمر أساسي في العديد من العمليات الحيوية خاصة بالنسبة لنمو حقيقيات النوى والدخول الفيروسي. يمكن أن تنشأ بروتينات الدمج من جينات تشفر لفيروسات مغلفة معدية، من فيروسات راجعة قديمة ضُمّنت في جينوم المضيف،[2] أو من جينات متواجدة ذاتيا في جينوم المضيف.[3] يمكن أن تؤثر تعديلات ما بعد الترجمة التي يقوم بها المضيف لبروتينات الدمج بشكل كبير على قدرتها في الدمج.[4]
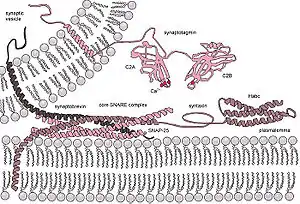
الدمج في حقيقيات النوى
تحتوي جينومات حقيقيات النوى على العديد من عائلات الجينات -من أصل ذاتي وفيروسي- التي تشفر نواتجا لها دور في إحداث اندماج الغشاء. في حين أن الخلايا الجسدية البالغة لا تخضع عادة لاندماج الغشاء تحت الظروف العادية، تتبع الجاميتات والخلايا الجنينية مسارات نمو لا تسبب اندماج الغشاء التلقائي [الإنجليزية] مثل: تشكل المشيمة، تشكل الأرومة الغاذية المخلوية والنمو العصبي. لمسارات الدمج دور كذلك في نمو الجهاز العضلي الهيكلي وأنسجة الجهاز العصبي. تعتمد عمليات دمج الحويصلة عند نقل النواقل العصبية على النشاط الإنزيمي لبروتينات الدمج.
من الأمثلة على بروتينات دمج الغشاء في حقيقيات النوى:
- عائلة SNARE: التي تعتبر مثالا على بروتينات دمج أصلية في حقيقيات النوى ولا تتواجد سوى لدى حقيقيات النوى والعتائق قريبة الصلة مثل عتائق أسغارد [الإنجليزية].[5] ومن الأمثلة عليها: بروتين فامب [الإنجليزية] (VAMP).
- بروتينات الفيروسات الراجعة: نشأت هذه البروتينات من بروتين جين env الخاص بالفيروسات الراجعة الداخلية وهي بروتينات دمج من الصنف 1 مستأنسة. ومن الأمثلة عليها بروتينات السينسيتين المسؤولة على بنُى المشيمة: سينسيتين 1 وسينسيتين 2. بروتين ERV3 لا فقد وظيفة دمج الأغشية.
- عائلة HAP2: بروتين HAP2 هو بروتين دمج فيروسي مستأنس من الصنف 2 يتواجد في مجموعة متنوعة من حقيقيات النوى مثل المقوسة الغوندية، رشاد أذن الفأر وذباب الفاكهة. هذا البروتين لازم من أجل اندماج الجاميت في هذه الكائنات.[6]
بروتينات دمج فيروسية
تتغلب الفيرسات المغلفة بسهولة على الحاجز الثرموديناميكي لدمج غشائي بلازما عبر تخزين طاقة حركية في بروتينات الدمج. يمكن التعبير عن بروتينات الدمج بشكل مستقل في سطح الخلية المضيفة وهو ما يمكن أن يقود إلى: (1) اندماج الخلية المصابة مع الخلايا المجاورة أو (2) إدراجه في فيروس متبرعم من الخلية المصابة وهو ما يؤدي إلى التحرر الكامل لغشاء البلازما من الخلية المضيفة. تقوم بعض بروتينات الدمج بعملية الدمج بمفردها بينما تحتاج مجموعة فرعية أخرى إلى التآثر مع عوامل خلوية. توجد أربع أقسام من بروتينات الدمج صُنفت حسب بنيتها وآلية الدمج.[7]
الصنف 1
يماثل الصنف الأول في بنيته الراصة الدموية الخاصة بفيروس الإنفلونزا. بعد الدمج، يملك الموقع النشط مثلوثا من لفات ملتفة للوالب ألفا. نطاق الارتباط غني بلوالب ألفا ويتواجد ببتيد الدمج الكاره للماء بالقرب من النهاية الأمينية. عادة ما يمكن التحكم في هيئة الدمج بواسطة الأس الهيدروجيني.[8][9]
الصنف 2
تهيمن صحائف بيتا على بروتينات الدمج من الصنف 2 وتتواجد المواقع المحفزة في منطقة اللب. مناطق الببتيد المطلوبة من أجل إحداث الدمج مكونة من لفات بين صحائف بيتا.[8][9]
الصنف 3
بروتينات الدمج من الصنف 3 مميزة عن بروتينات الصنف 1 و2. وتتكون عادة من خمس نطاقات بنيوية يتواجد فيها النطاقان 1 و2 ناحية النهاية الكربوكسيلية وعادة ما يحتويان على صحائف بيتا أكثر، النطاقات 2-5 أقرب إلى جانب النهاية الأمينية وأغنى بلوالب ألفا. في هيئة قبل الاندماج تحتضن النطاقات الأخيرة النطاق 1 وتحميه (النطاق 1 محمي بالنطاق 2 الذي يحتضنه النطاق 3 الذي بدوره محمي بواسطة النطاق 4). يحتوي النطاق 1 على الموقع التحفيزي من أجل دمج الغشاء.[8][9]
الصنف 4
الصنف الرابع من بروتينات الدمج معروف باسم البروتينات عبر الغشائية الصغيرة المرتبطة بالدمج (FAST)، وهي أصغر أنواع بروتينات الدمج. تتواجد في الفيروسات الراجعة وهي فيروسات غير مغلفة ومختصة بدمج خلية-خلية بدل فيروس-خلية وتشكل ملتحمات خلوية. وهي النوع الوحيد من بروتينات دمج الغشاء التي وُجدت في الفيروسات غير المغلفة.[10][11]
أمثلة
| بروتين الدمج | الاختصار | الصنف | عائلة الفيروس | أمثلة فيروسية | مراجع |
|---|---|---|---|---|---|
| شوكة فيروس كورونا | S | 1 | فيروسات تاجية | سارس-كوف، سارس-كوف-2 | [12][13] |
| البروتين السكري لفيروس إيبولا | GP | 1 | فيروسات خيطية | فيروس إيبولا السودان، الزائير، فيروس ماربورغ | [7][14] |
| البروتين السكري gp41 | Gp41 | 1 | فيروس راجع | فيروس العوز المناعي البشري | [7][14] |
| راصة دموية (إنفلونزا) | H ،HA ،HN | 1 | فيروس الإنفلونزا، فيروسات مخاطية | فيروس الإنفلونزا، فيروس النكاف، فيروس الحصبة | [7][14] |
| بروتين غلاف فيروس ألفا E1 | E1 | 2 | فيروسات طخائية | فيروس غابة سيمليكي | [7][14] |
| بروتين غشاء فيروس المصفر | E | 2 | فيروسات مصفرة | فيروس الضنك، فيروس غرب النيل | [7][14] |
| البروتين السكري B لفيروس الهربس | gB | 3 | فيروسات هربسية | فيروس الهربس البسيط | [7][15] |
| بروتين G لفيروس التهاب الفم الحويصلي (VSV G) | G | 3 | فيروسات ربدية | فيروس داء الكلب، فيروس التهاب الفم الحويصلي | [7][15] |
| البروتين عبر الغشائي الصغير المرتبط بالدمج | FAST | 4 | فيروسات تنفسية معوية يتيمة | فيروس ريو المستقيم الطيري [الإنجليزية] | [7][11] |
مراجع
- "Subneuronal processing of information by solitary waves and stochastic processes"، Nano and Molecular Electronics Handbook، Nano and Microengineering Series، CRC Press، 2007، ص. 17–1–17–41، doi:10.1201/9781420008142.ch17 (غير نشط 31 أكتوبر 2021)، ISBN 978-0-8493-8528-5، مؤرشف من الأصل في 16 يناير 2016، اطلع عليه بتاريخ 30 يناير 2022.
{{استشهاد بكتاب}}: صيانة CS1: وصلة دوي غير نشطة منذ 2021 (link) - Classification of viral fusion proteins in قاعدة بيانات تصنيف الناقل database نسخة محفوظة 2020-08-09 على موقع واي باك مشين.
- "The formation of syncytia within the visceral musculature of the Drosophila midgut is dependent on duf, sns and mbc"، Mechanisms of Development، 110 (1–2): 85–96، يناير 2002، doi:10.1016/S0925-4773(01)00567-6، PMID 11744371، مؤرشف من الأصل في 30 يناير 2022.
- "Addicted to sugar: roles of glycans in the order Mononegavirales"، Glycobiology، 29 (1): 2–21، يناير 2019، doi:10.1093/glycob/cwy053، PMC 6291800، PMID 29878112.
- "Prototypic SNARE Proteins Are Encoded in the Genomes of Heimdallarchaeota, Potentially Bridging the Gap between the Prokaryotes and Eukaryotes"، Current Biology، 30 (13): 2468–2480.e5، يوليو 2020، doi:10.1016/j.cub.2020.04.060، PMID 32442459.
- "The Ancient Gamete Fusogen HAP2 Is a Eukaryotic Class II Fusion Protein"، Cell، 168 (5): 904–915.e10، فبراير 2017، doi:10.1016/j.cell.2017.01.024، PMC 5332557، PMID 28235200.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - Podbilewicz, Benjamin (11 أكتوبر 2014)، "Virus and Cell Fusion Mechanisms"، Annual Review of Cell and Developmental Biology، 30 (1): 111–139، doi:10.1146/annurev-cellbio-101512-122422.
- "Class III viral membrane fusion proteins"، Current Opinion in Structural Biology، 19 (2): 189–96، أبريل 2009، doi:10.1016/j.sbi.2009.02.012، PMC 3076093، PMID 19356922.
- "Structures and mechanisms of viral membrane fusion proteins: multiple variations on a common theme"، Critical Reviews in Biochemistry and Molecular Biology، 43 (3): 189–219، 2008، doi:10.1080/10409230802058320، PMC 2649671، PMID 18568847.
- Shmulevitz, Maya؛ Duncan, Roy (01 مارس 2000)، "A new class of fusion-associated small transmembrane (FAST) proteins encoded by the non-enveloped fusogenic reoviruses"، The EMBO Journal، 19 (5): 902–912، doi:10.1093/emboj/19.5.902، PMC 305630.
- Ciechonska, Marta؛ Duncan, Roy (ديسمبر 2014)، "Reovirus FAST proteins: virus-encoded cellular fusogens"، Trends in Microbiology، 22 (12): 715–724، doi:10.1016/j.tim.2014.08.005.
- Li, Fang (29 سبتمبر 2016)، "Structure, Function, and Evolution of Coronavirus Spike Proteins"، Annual Review of Virology، 3 (1): 237–261، doi:10.1146/annurev-virology-110615-042301، PMC 5457962.
- Zhu, Chaogeng؛ He, Guiyun؛ Yin, Qinqin؛ Zeng, Lin؛ Ye, Xiangli؛ Shi, Yongzhong؛ Xu, Wei (أكتوبر 2021)، "Molecular biology of the SARs‐CoV‐2 spike protein: A review of current knowledge"، Journal of Medical Virology، 93 (10): 5729–5741، doi:10.1002/jmv.27132، PMC 8427004، PMID 34125455.
- White, Judith M.؛ Whittaker, Gary R. (يونيو 2016)، "Fusion of Enveloped Viruses in Endosomes"، Traffic، 17 (6): 593–614، doi:10.1111/tra.12389، PMC 4866878.
- Baquero, Eduard؛ Albertini, Aurélie AV؛ Gaudin, Yves (أغسطس 2015)، "Recent mechanistic and structural insights on class III viral fusion glycoproteins"، Current Opinion in Structural Biology، 33: 52–60، doi:10.1016/j.sbi.2015.07.011.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية