بوليمراز الدنا
بوليمراز الدنا (بالإنجليزية: DNA polymerase) هو إنزيم مساعد في عملية تناسخ الحمض النووي الريبي منقوص الأكسجين حيث يقوم بتحفيز عملية بلمرة النيوكليوتيدات، الوحدة البنائية لحمض الدنا. يعمل بوليميراز الدنا ضمن مجموعة من الإنزيمات المحفزة لعملية تضاعف الحمض النووي، حيث تعمل هذه الإنزيمات جميعًا لتصنيع نسخة من الحمض النووي مطابقة لسلسلة الحمض النووي الأصلية.[1][2] يكمن دور البوليمراز في قراءة السلسلة الأساسية وترتيب النيوكليوتيدات في صورة مطابقة للسلسلة الأصلية.[3][4] في كل مرة تنقسم فيها الخلية، يعمل بوليميراز الدنا مع الإنزيمات الأخرى لاستنساخ الحمض النووي وتزويد الخلايا الجديدة بنسخ من الحمض النووي مطابقة للحمض النووي الموجود في الخلية الأم. وبهذه الطريقة، تنقل المعلومات الجينية من جيل لآخر.
| بوليميراز الدنا | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
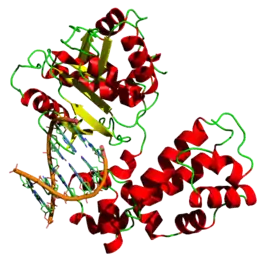 رسم ثلاثي الأبعاد لإنزيم البوليميراز عند الإنسان | |||||||||||
| أرقام التعريف | |||||||||||
| رقم التصنيف الإنزيمي | 2.7.7.7 | ||||||||||
| رقم التسجيل CAS | 9012-90-2 | ||||||||||
| قواعد البيانات | |||||||||||
| قاعدة بيانات الإنزيم | راجع IntEnz | ||||||||||
| قاعدة بيانات براونشفايغ | راجع BRENDA | ||||||||||
| إكسباسي | راجع NiceZyme | ||||||||||
| موسوعة كيوتو | راجع KEGG | ||||||||||
| ميتاسيك | المسار الأيضي | ||||||||||
| بريام | ملف التعريف | ||||||||||
| تركيب بنك بيانات البروتين | RCSB PDB PDBe PDBsum | ||||||||||
| الأونتولوجيا الجينية | AmiGO / EGO | ||||||||||
| |||||||||||
و هو يستخدم أيون المغنيسيوم كعامل حفاز.
التاريخ
في 1956م، اكتشف عالم الكيمياء الحيوية آرثر كورنبرغ وزملائه بوليميراز الدنا الأول في بكتيريا الإشريكية القولونية. استطاع الفريق شرح خطوات تضاعف الحمض النووي عن طريق إنزيم البوليميراز. ونتيجة لهذا الاكتشاف، نال كورنبرغ جائزة نوبل في الطب أو علم وظائف الأعضاء عام 1959م.[5] أما بوليميراز الدنا الثاني فقد اكتشف على يد عالم الكيمياء الحيوية توماس كونبرغ (ابن آرثر كورنبرغ) ومالكوم جيفتر في عام 1970م، أثناء محاولتهما لكشف المزيد عن دور البوليميراز الأول في تضاعف الحمض النووي عند بكتيريا الإشريكية القولونية.[6] لاحقًا، اكتشفت ثلاثة أنواع أخرى من البوليميراز في بكتيريا الإشريكية القولونية، الثالث في السبيعينيات، والرابع والخامس في عام 1999م.[7]
وظيفته
قبل أن تبدأ عملية التضاعف، يقوم إنزيم الهيليكاز بحل سلسلتي الدنا عن طريق كسر الروابط الهيدروجينية بين النيوكليوتيدات. ينتج عن هذه العملية فك السلسلتين المترابطتين للحمض النووي؛ مما يمكن باقي الإنزيمات من العمل على السلسلتين المنفردتين. يعمل البلوليميراز على قراءة كل سلسلة على حدة، وإضافة النيوكليوتيدات واحدة تلو الأخرى إلى النهاية 3' لسلسلة الحمض النووي. تُضاف النيوكليوتيدات وفق ترتيب محدد مبني على نوع القاعدة النيتروجينية في النيوكليوتيد؛ حيث يُضاف السايتوسين للجوانين، والثايمين للأدينين. ويمكن لبوليميراز الدنا بوليمريز إضافة نيوكليوتيدة جديدة إلى النهاية 3' من السلسلة الجديدة، مما يؤدي إلى استطالة السلسلة في اتجاه واحد هو من 5' إلى 3'. حتى الآن، لا يوجد إنزيم بوليميراز واحد قادر على تكوين سلسلة حمض نووي من العدم؛ إذ يتطلب عمل هذا الإنزيم دائمًا وجود نهاية 3' مسبقًا. وتسمى هذه الجزيئات التي يقوم البوليميراز بإضافة النيوكليوتيدات إليها بالمشرعات أو البادئات. وقد تتكون هذه المشرّعات من حمض الدنا، أو حمض الرنا، أو الحمضين معًا. أثناْ تضاعف الحمض النووي، تتكون القاعدتان الأوليتان دائمًا من حمض الرنا، ويُصنّعان بواسطة إنزيم البريميز. كما يتطلب عمل البوليميريز حل سلسلتي الدنا، عن طريق إنزيم الهيليكاز وإنزيم التوبوإيزوميراز، في آلية تضاعف تعرف باسم التضاعف نصف المحافظ.[8]
من المهم ملاحظة أنه بينما يكوّن البوليميراز سلسلة متجهة من 5' إلى 3'، فإنه يتحرك باتجاه معاكس على السلسلة الأصلية؛ أي أنه يمشي باتجاه 3' إلى 5'. هذا الاختلاف يسمح بإنشاء سلسلتين من الدنا بترتيب يسمى بالتوازي المتضاد.[9]
في بعض الأحيان، قد يشوب عمل الإنزيم بعض الأخطاء؛ يحدث هذا بمعدل خطأ واحد لكل مليار نيوكليوتيدة مركبة. تملك بعض أنواع البوليميراز خاصية تصحيح الأخطاء. حين يتم التعرف على خطأ في تركيبة النيوكليوتيدات، يتراجع الإنزيم خطوة تلو أخري للوراء. تسمح خاصية النوكلياز الخارجية لدى هذه الإنزيمات بإزالة النيوكليوتيد الخطأ ثم إعادى تركيب النيوكليوتيدات الصحيحة في مكانها. تسمى هذه الظاهرة بالتصحيح؛ وهي ضرورية للحفاظ على التطابق بين السلسلة الجديدة والسلسلة الأصلية.[10] للدقة أهمية كبرى في عملية تضاعف الحمض النووي، فالأخطاء التركيبية قد تُنتج بروتينات مشوهة تقود إلى تكوين الأورام السرطانية.
البناء
تتمتع بوليميرات الحمض النووي المعروفة ببُنية محمية للغاية، مما يعني أن وحداتها الفرعية التحفيزية الشاملة تختلف قليلاً جدًا من نوع إلى نوع، بغض النظر عن هياكل مجالها. تشير الهياكل المحفوظة عادةً إلى وظائف الخلية المهمة التي لا يمكن الاستغناء عنها، والتي توفر صيانتها مزايا تطورية. يمكن وصف الشكل بأنه يشبه اليد اليمنى مع مجالات الإبهام والإصبع وراحة اليد (Palm). يبدو أن مجال راحة اليد النخيل يعمل في تحفيز نقل مجموعات الفسفوريل في تفاعل نقل الفسفوريل. يرتبط الحمض النووي براحة اليد عندما يكون الإنزيم نشطًا. يُعتقد أن هذا التفاعل يُحفَّز بواسطة آلية ثنائية المعدن أيون (Two-metal-ion mechanism). يعمل مجال الإصبع على ربط النيوكليوسيد ثلاثي الفوسفات بقاعدة القالب. يلعب مجال الإبهام دورًا محتملاً في العملية، والانتقال، وتحديد موضع الحمض النووي.[11]
العملية
يرجع التحفيز السريع لبوليميراز الحمض النووي إلى طبيعته المعالجة. العملية هي سمة من سمات الإنزيمات التي تعمل على ركائز بوليمرية. في حالة بوليميريز الحمض النووي، تشير درجة العملية إلى متوسط عدد النيوكليوتيدات المُضافة في كل مرة يربط الإنزيم قالبًا. يتطلب متوسط بوليميريز الحمض النووي حوالي ثانية واحدة لتحديد وربط التمهيدي/ تقاطع القالب. بمجرد أن يتم ربطها، تضيف بوليميراز الدنا غير العملي نيوكليوتيدات بمعدل نيوكليوتيد واحد في الثانية. درجة العملية تتناسب طرديا مع معدل تصنيع الحمض النووي. حُدِّد معدل تصنيع الحمض النووي في الخلية الحية لأول مرة على أنه معدل استطالة الحمض النووي للعاثية الحمض النووي T4 في العدوى بالبكتيريا القولونية. خلال فترة زيادة الحمض النووي الأسي عند 37 درجة مئوية، كان المعدل 749 نيوكليوتيد في الثانية.
تسمح قدرة بوليميراز الحمض النووي على الانزلاق على طول قالب الحمض النووي بزيادة المعالجة. هناك زيادة كبيرة في المعالجة عند شوكة النسخ المتماثل. تُسَهَّل هذه الزيادة من خلال ارتباط بوليميريز الحمض النووي بالبروتينات المعروفة باسم مشبك الحمض النووي المنزلق. المشابك عبارة عن وحدات فرعية بروتينية متعددة مرتبطة بشكل حلقة. باستخدام التحلل المائي لأدينوسين ثلاثي الفوسفات (ATP)، تفتح فئة من البروتينات تُعرف باسم بروتينات تحميل المشبك المنزلق الهيكل الدائري لمشابك الحمض النووي المنزلق مما يسمح بالارتباط والإفراج عن حبلا DNA. يمنع تفاعل البروتين والبروتين مع المشبك بوليميريز الحمض النووي من الانتشار من قالب الحمض النووي، وبالتالي ضمان أن الإنزيم يربط نفس التقاطع التمهيدي/ القالب ويستمر في التكرار.[12][13]
انظر أيضًا
مراجع
- "معلومات عن بوليميراز الدي إن إيه على موقع chem.nlm.nih.gov"، chem.nlm.nih.gov، مؤرشف من الأصل في 6 أغسطس 2020.
- "معلومات عن بوليميراز الدي إن إيه على موقع echa.europa.eu"، echa.europa.eu، مؤرشف من الأصل في 17 نوفمبر 2019.
- Bollum, F. J. (01 يناير 1963)، "Studies on the Nature of Calf Thymus DNA-Polymerase Products"، Cold Spring Harbor Symposia on Quantitative Biology، 28 (0): 21–26، doi:10.1101/sqb.1963.028.01.008، ISSN 0091-7451، مؤرشف من الأصل في 8 يناير 2021.
- SZULMAJSTER, J. (01 سبتمبر 1973)، "Recent Studies on the Deoxyribonucleic Acid-Dependent Ribonucleic Acid Polymerase from Vegetative Cells and Spores of Bacillus subtilis"، Biochemical Society Transactions، 1 (5): 1021–1022، doi:10.1042/bst0011021، ISSN 0300-5127، مؤرشف من الأصل في 8 يناير 2021.
- Jan؛ Ringertz, Nils (2001-08)، The Nobel Prize، CO-PUBLISHED WITH IMPERIAL COLLEGE PRESS، ص. 111–134، ISBN 978-981-02-4664-8، مؤرشف من الأصل في 8 يناير 2021.
{{استشهاد بكتاب}}: تحقق من التاريخ في:|تاريخ=(مساعدة) - "The ARule and Deletion Formation During Abasic and Oxidized Abasic Site Bypass by DNA Polymerase" (PDF)، dx.doi.org، مؤرشف من الأصل في 8 يناير 2021، اطلع عليه بتاريخ 08 يناير 2021.
- Lehninger principles of biochemistry (ط. 6th ed)، New York: W.H. Freeman and Company، 2013، ISBN 978-1-4292-3414-6، OCLC 824794893، مؤرشف من الأصل في 3 أغسطس 2020.
{{استشهاد بكتاب}}:|edition=has extra text (مساعدة) - An introduction to genetic analysis (ط. 7th ed)، New York: W.H. Freeman، 2000، ISBN 0-7167-3520-2، OCLC 42049331، مؤرشف من الأصل في 8 يناير 2021.
{{استشهاد بكتاب}}:|edition=has extra text (مساعدة) - Burge, Sarah؛ Parkinson, Gary N.؛ Hazel, Pascale؛ Todd, Alan K.؛ Neidle, Stephen (2006-11)، "Quadruplex DNA: sequence, topology and structure"، Nucleic Acids Research، 34 (19): 5402–5415، doi:10.1093/nar/gkl655، ISSN 0305-1048، PMID 17012276، مؤرشف من الأصل في 8 مايو 2020.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة) - "Mary Elizabeth Garrett"، 2008، doi:10.1353/book.3515، مؤرشف من الأصل في 8 يناير 2021.
{{استشهاد بدورية محكمة}}: Cite journal requires|journal=(مساعدة) - Steitz, T. A. (18 يونيو 1999)، "DNA polymerases: structural diversity and common mechanisms"، The Journal of Biological Chemistry، 274 (25): 17395–17398، doi:10.1074/jbc.274.25.17395، ISSN 0021-9258، PMID 10364165، مؤرشف من الأصل في 20 أغسطس 2020.
- Molecular biology of the gene (ط. 6th ed)، San Francisco: Pearson/Benjamin Cummings، 2008، ISBN 978-0-8053-9592-1، OCLC 173182652، مؤرشف من الأصل في 15 ديسمبر 2018.
{{استشهاد بكتاب}}:|edition=has extra text (مساعدة) - McCarthy, D.؛ Minner, C.؛ Bernstein, H.؛ Bernstein, C. (05 أكتوبر 1976)، "DNA elongation rates and growing point distributions of wild-type phage T4 and a DNA-delay amber mutant"، Journal of Molecular Biology، 106 (4): 963–981، doi:10.1016/0022-2836(76)90346-6، ISSN 0022-2836، PMID 789903، مؤرشف من الأصل في 23 نوفمبر 2020.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة طب
بوابة طب بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية