تطور الجينوم
تطور الجينوم هي العملية التي يتغير الجينوم بوساطتها من حيث البنية (التسلسل) أو الحجم مع الزمن. تتضمن دراسة تطور الجينوم مجالات متعددة كالتحليل البنيوي للجينوم ودراسة الطفيليات الجينومية ودراسة الجينات وتضاعفات الجينوم القديمة وتعدد الصيغ الكروموسومية وعلم الجينوم المقارن.

نبذة تاريخية
بدأت سلسلة نماذج الجينوم الأولى في أواخر السبعينيات. ومنذ ذلك الحين، بدأ العلماء باستخدام علم الجينوم المقارن لدراسة أوجه التشابه والاختلاف بين النماذج المختلفة. تقدم تسلسل الجينوم بمرور الوقت ليشمل الكثير من الجينومات المعقدة، بما في ذلك التسلسل النهائي للجينوم البشري بأكمله في عام 2001. سمحت المقارنة بين جينوم الأقارب والأسلاف البعيدين بتوضيح أوجه التشابه والاختلاف وبينت الآليات التي ستقود تطور الجينوم.
جينومات بدائية وحقيقية النواة
بدائيات النوى
تؤثر آليتان رئيسيتان على تطور جينوم بدائيات النواة هما: الطفرات ونقل الجينات الأفقي.[1] يعتبر التكاثر الجنسي الآلية الرئيسية الثالثة في حقيقيات النوى، ولكنه يؤثر أيضًا على البكتيريا. قد تكتسب بدائيات النوى مادة وراثية جديدة من خلال عملية الاقتران البكتيري؛ حيث تنتقل البلازميدات والكروموسومات كاملة. يعتبر انتقال المقاومة على المضادات الحيوية عبر البلازميدات من الأمثلة الشائعة عن الأمر.[2] يتطور الجينوم أيضًا عن طريق آلية التنبيغ، وفيه تقوم العاثيات بإدخال حمض نووي ريبوزي منقوص الأكسجين جديد إلى جينوم بكتيري ما. تتمثل الآلية الرئيسية للتفاعل الجنسي بالتحول الجيني الطبيعي الذي يتضمن نقل الحمض النووي من خلية بدائية النواة إلى أخرى من خلال وسيط. يعتبر التحول أحد الأنماط الشائعة لانتقال الحمض النووي، وقد وثق الباحثون أهلية 67 نوع بدائي النواة على الأقل للتحول.[3]
خضع تطور الجينوم في البكتيريا للدراسة ويعتبر أحد الأمور المفهومة بشكل جيد بسبب توفر آلاف الجينومات البكتيرية المسلسلة بشكل كامل. قد تؤدي التغييرات الجينية إلى زيادة أو نقصان التعقيد الجيني بسبب الانتقاء المبسط المنقي.[4] بشكل عام، طورت البكتيريا التي تعيش في بحرية في الطبيعة جينوم أكبر يحوي جينات أكثر لتتمكن من التكيف بسهولة أكبر مع الظروف البيئية المتغيرة. على النقيض من ذلك، قللت معظم البكتيريا الطفيلية من الجينوم بسبب حصولها على العديد من المغذيات من المضيف؛ أي لا يحتاج جينومهم إلى ترميز الإنزيمات المنتجة لهذه العناصر الغذائية.[5]
* تحتوي جينات الإشريكية القولونية على الإكسونات فقط في معظم الحالات، وقد تحتوي على كميات صغيرة من الإنترونات ذاتية التضفير في بعض الأحيان (المجموعة الثانية).[6]
| الصفة | جينوم الإشريكية القولونية | الجينوم البشري |
| حجم الجينوم (أزواج قاعدية) | 4.6 ميغا قاعدة | 3.2 جيغا قاعدة |
| هيكل الجينوم | حلقي | خطي |
| عدد الكروموسومات | 1 | 46 |
| وجود البلازميدات | نعم | لا |
| وجود هستونات | لا | نعم |
| فصل الحمض النووي في النواة | لا | نعم |
| عدد الجينات | 4288 | 20000 |
| وجود الإنترونات | لا * | نعم |
| متوسط حجم الجين | 700 زوج قاعدي | 27000 زوج قاعدي |
حقيقيات النوى
تكون جينومات حقيقيات النوى أكبر من بدائيات النوى عمومًا. يبلغ طول جينوم الإشريكية القولونية 4.6 ميغا قاعدة تقريبًا،[7] بينما يكون الجينوم البشري أكبر بكثير بحجم 3.2 جيغا قاعدة تقريبًا.[8] يأخذ جينوم حقيقيات النوى الهيكل الخطي وقد يتألف من عدة كروموسومات تتوضع داخل نواة الخلية. تزيل عملية تضفير الحمض النووي الريبوزي –قبل ترجمة البروتين- الإنترونات، وهي أجزاء غير فعالة من الجين لا توجد في معظم بدائيات النوى. يتطور جينوم حقيقيات النوى بمرور الوقت عبر عدة آليات؛ كالتكاثر الجنسي الذي يوفر تنوع جيني كبير للنسل مقارنةً بالتكاثر المعتاد لدى بدائيات النوى؛ ففيه يكون النسل استنساخ وراثي بحت للخلية الأم.
حجم الجينوم
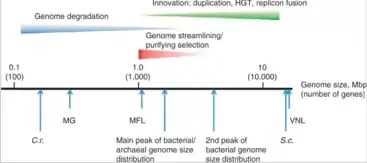
يقاس حجم الجينوم عادةً بالأزواج القاعدية (قواعد الحمض النووي الريبوزي منقوص الأكسجين أحادي السلسلة أو الحمض النووي الريبوزي). يمكن قياس حجم الجينوم بواسطة القيمة سي أيضًا. أظهرت الأبحاث التي أجريت على جينومات بدائيات النوى وجود علاقة ترابط طردية واضحة بين القيمة سي لبدائيات النوى وكمية الجينات التي يتكون منها الجينوم.[9] يشير ذلك إلى أهمية عدد الجينات باعتباره العامل الرئيسي المؤثر على حجم جينوم بدائية النواة. لا يلاحظ الأمر في حقيقيات النوى؛ أي لا يتناسب عدد الجينات لديهم مع حجم الجينوم. وبعبارة أخرى، يكون حجم الجينوم أكبر بكثير مما هو متوقع عند النظر إلى العدد الإجمالي للجينات المشفرة للبروتين.[10]
يزداد حجم الجينوم عبر آليات التكرار والغرز وتعدد الصيغ الصبغية. قد تزيد عملية إعادة التركيب الجيني من حجم الجينوم أو تنقصه. يؤدي الحذف الجيني إلى إنقاص حجم الجينوم. تعتبر المتفطرات الجذامية -العامل المسبب للجذام- أحد الأمثلة الشهيرة على التقلص الجيني؛ إذ فقدت المتفطرات الجذامية العديد من الجينات الفعالة سابقًا بسبب تشكل الجينات الكاذبة.[11] يشاهد ذلك بوضوح لدى أقرب أسلافها المتفطرات السلية.[12] تعيش المتفطرات الجذامية وتتكاثر داخل عائل، ولهذا لا تحتاج إلى العديد من الجينات التي كانت تحملها في السابق لتمكنها من النجاة والاستمرار خارج العائل. وهكذا، فقدت هذه الجينات وظيفتها بمرور الوقت عبر عدة آليات حولتها لجينات كاذبة، وتعتبر الطفرات مثال على تلك الآليات. يستفيد الكائن الحي من التخلص من الجينات غير الأساسية؛ بسبب ارتفاع سرعة تضاعف الحمض النووي وانخفاض الطاقة المستهلكة في هذه العملية.[13]
تعتبر العوامل الممرضة الخيطية التي تصيب النبات أمثلة واضحة على ازدياد حجم الجينوم بمرور الوقت. نمى جينوم تلك العوامل الممرضة بشكل كبير على مر السنين بسبب التكرار. احتوت المناطق الغنية بالتكرارات على جينات ترمز بروتينات التفاعل مع المضيف. أدت إضافة التكرارات إلى ارتفاع احتمال تطوير عوامل فوعة جديدة عبر حدوث الطفرات وأشكال إعادة التركيب الجيني الأخرى. بهذه الحالة، استفادت العوامل الممرضة الخيطية من ازدياد حجم الجينوم.[14]

الانتواع
تعتبر الطريقة التي يتغير فيها الجينوم ليخلق أنواع جديدة سؤالًا رئيسيًا في علم الأحياء التطوري. يتطلب الانتواع حدوث تغيرات في السلوك أو الشكل أو الفيزيولوجيا أو الأيض (بالإفراد أو الجمع). بدأت دراسة تطور الجينوم خلال عملية الانتواع للدراسة منذ وقت قصير بالتزامن مع توفر تقنيات السلسلة الحديثة. على سبيل المثال، تختلف أسماك البلطي في البحيرات الأفريقية في الشكل والسلوك. كشفت جينومات 5 أنواع تغير التسلسل ونمط التعبير الجيني خلال فترة زمنية قصيرة نسبيًا (100000 إلى عدة ملايين من السنين). اكتسب 20٪ من أزواج الجينات المكررة نمط تعبير نسيجي جديد تمامًا، وهذا يقترح حصول هذه الجينات على وظائف جديدة أيضًا. يعتمد اختلاف التعبير الجيني على اختلاف تسلسلات تنظيمية قصيرة، وهذا يشير إلى أن حدوث الانتواع لا يتطلب سوى عدد قليل نسبيًا من الطفرات. أظهر جينوم البلطيات أيضًا ارتفاع في المعدلات التطورية للحمض الريبوزي النووي الميكروي الذي يساهم في التعبير الجيني.[15][16]
مراجع
- Toussaint A, Chandler M (2012)، "Prokaryote genome fluidity: toward a system approach of the mobilome"، Bacterial Molecular Networks، Methods in Molecular Biology، ج. 804، ص. 57–80، doi:10.1007/978-1-61779-361-5_4، ISBN 978-1-61779-360-8، PMID 22144148.
- Ruiz J, Pons MJ, Gomes C (سبتمبر 2012)، "Transferable mechanisms of quinolone resistance"، International Journal of Antimicrobial Agents، 40 (3): 196–203، doi:10.1016/j.ijantimicag.2012.02.011، PMID 22831841.
- Johnsborg O, Eldholm V, Håvarstein LS. Natural genetic transformation: prevalence, mechanisms and function. Res Microbiol. 2007 Dec;158(10):767-78. doi: 10.1016/j.resmic.2007.09.004. Epub 2007 Oct 2. PMID 17997281
- Koonin EV, Wolf YI (ديسمبر 2008)، "Genomics of bacteria and archaea: the emerging dynamic view of the prokaryotic world"، Nucleic Acids Research، 36 (21): 6688–719، doi:10.1093/nar/gkn668، PMC 2588523، PMID 18948295.
- Tortora, Gerard J. (2015)، Microbiology: An Introduction، ISBN 978-0321929150.
- Dai L, Zimmerly S (أكتوبر 2002)، "The dispersal of five group II introns among natural populations of Escherichia coli"، RNA، 8 (10): 1294–307، doi:10.1017/S1355838202023014، PMC 1370338، PMID 12403467.
- Blattner FR, Plunkett G, Bloch CA, Perna NT, Burland V, Riley M, وآخرون (سبتمبر 1997)، "The complete genome sequence of Escherichia coli K-12"، Science، 277 (5331): 1453–62، doi:10.1126/science.277.5331.1453، PMID 9278503.
- International Human Genome Sequencing Consortium (أكتوبر 2004)، "Finishing the euchromatic sequence of the human genome"، Nature، 431 (7011): 931–45، Bibcode:2004Natur.431..931H، doi:10.1038/nature03001، PMID 15496913.
- Gregory TR (فبراير 2001)، "Coincidence, coevolution, or causation? DNA content, cell size, and the C-value enigma"، Biological Reviews of the Cambridge Philosophical Society، 76 (1): 65–101، doi:10.1017/S1464793100005595، PMID 11325054.
- Gregory TR (يناير 2002)، "A bird's-eye view of the C-value enigma: genome size, cell size, and metabolic rate in the class aves"، Evolution; International Journal of Organic Evolution، 56 (1): 121–30، doi:10.1111/j.0014-3820.2002.tb00854.x، PMID 11913657.
- Singh P, Cole ST (يناير 2011)، "Mycobacterium leprae: genes, pseudogenes and genetic diversity"، Future Microbiology، 6 (1): 57–71، doi:10.2217/fmb.10.153، PMC 3076554، PMID 21162636.
- Eiglmeier K, Parkhill J, Honoré N, Garnier T, Tekaia F, Telenti A, وآخرون (ديسمبر 2001)، "The decaying genome of Mycobacterium leprae"، Leprosy Review، 72 (4): 387–98، doi:10.5935/0305-7518.20010054، PMID 11826475.
- Rosengarten R, Citti C, Glew M, Lischewski A, Droesse M, Much P, وآخرون (مارس 2000)، "Host-pathogen interactions in mycoplasma pathogenesis: virulence and survival strategies of minimalist prokaryotes"، International Journal of Medical Microbiology، 290 (1): 15–25، doi:10.1016/S1438-4221(00)80099-5، PMID 11043978.
- Raffaele S, Kamoun S (مايو 2012)، "Genome evolution in filamentous plant pathogens: why bigger can be better"، Nature Reviews. Microbiology، 10 (6): 417–30، doi:10.1038/nrmicro2790، PMID 22565130، S2CID 6169712.
- Jiggins CD (سبتمبر 2014)، "Evolutionary biology: Radiating genomes"، Nature، 513 (7518): 318–9، Bibcode:2014Natur.513..318J، doi:10.1038/nature13742، PMID 25186726.
- Brawand D, Wagner CE, Li YI, Malinsky M, Keller I, Fan S, وآخرون (سبتمبر 2014)، "The genomic substrate for adaptive radiation in African cichlid fish"، Nature، 513 (7518): 375–381، Bibcode:2014Natur.513..375B، doi:10.1038/nature13726، PMC 4353498، PMID 25186727.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي