تطور موجه
التطوُّر الموجَّه Directed evolution، هو عملية تستخدم في هندسة البروتينات وتغييرها بشكلٍ يحاكي عملية الانتقاء الطبيعي لتوجيهها نحو هدف محدَّد ومرسوم مسبقاً[1]، يشمل التطوُّر الموجَّه عدة خطوات تتمثَّل في إخضاع الجين لعدة جولات مُتكرِّرة من الطفرات والانتقاء والتضخيم، ويمكن القيام بهذه العمليَّة في الكائن الحي أو في المختبر، يستخدم التطوُّر الموجَّه لدراسة مبادئ التطور الأساسيَّة ضمن بيئة مختبريَّة محكومة أو مُسيطر عليها.
تاريخ
يرجع أصل التطوُّر الموجَّه إلى ستينات القرن الماضي[2]، عندما بدأ تطوير جزيئات RNA المُضخَّمة في التجارب المخبريَّة[3]، وامتدَّ هذا المفهوم لاحقاً ليشمل تطوُّر البروتينات الذي يحدث خلال تطوُّر الجراثيم تحت تأثير الضغوط الانتقائيَّة.[4]
سمحت تقنيَّات الهندسة الوراثية التي تمَّ تطويرها في ثمانينات القرن الماضي بإمكانية إجراء تعديلات دقيقة على بروتين واحد فقط [5]، ومكَّن هذا من اختيار بروتين واحد والقيام بالتعديلات عليه ولكنَّه لم يكن متوافقاً مع اختيار النشاط التحفيزي للأنزيمات[6]، لاحقاً وفي التسعينات تمَّ تطوير إمكانية التداخل على الأنزيمات واستخدمت هذه التقنيَّة على نطاق علمي واسع[7]، بعدها توسَّعت تقنيَّات التطوُّر الموجَّه بشكلٍ كبير وسريع مع ظهور طرق جديدة لخلق سلاسل من التغيُّرات الجينيَّة ومراقبة تأثيراتها[2][8]، أخيراً وفي عام 2018 تمَّ منح جائزة نوبل في الكيمياء لفرانسيس أرنولد وجورج سميث وغريغوري وينتر عن أبحاثهم في مجال تقنيَّات التطوُّر الموجَّه.[9]
المبادئ
يُشبه التطوُّر الموجَّه تسلُّق تلَّة أو مرتفع ما، حيث يمثِّل المرتفع الميِّزة أو الخاصيَّة المرغوب بها ويتمُّ تكرار التغيُّرات أو الطفرات حتى يتمُّ الوصول إلى التأثير المطلوب والذي هو أعلى ارتفاع أو قمة ممكنة، إنَّ التطوُّر المباشر هو محاكاة لدورة التطور الطبيعي ولكن ضمن بيئة مختبريَّة يتمُّ التحكُّم بها، يتطلُّب التطوُّر حدوث ثلاثة أشياء: حدوث التغيُّر بين النسخ المتماثلة، هذا الاختلاف سيؤدي بدوره إلى اختلافات عديدة في التلاؤم مع البيئة أو الظروف المحيطة وفي النهايات هذا التنوُّع سيتمُّ توريثه للأجيال القادمة [10]، تتكرَّر هذه الخطوات عادةً باستخدام أفضل صيغة من جولة واحدة كقالب واحد لتحقيق التحسينات التدريجيَّة، وترتبط احتماليَّة نجاح التطوُّر الموجَّه ارتباطاً مباشراً بحجم هذه التغيُّرات الإجمالي.[10]
توليد التباين
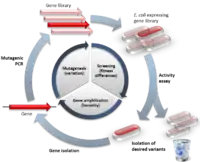
الخطوة الأولى في تنفيذ دورة من التطوُّر الموجَّه هي إنشاء «مكتبة» من الجينات المختلفة، بحيث يكون مجال التسلسل العشوائي شاسع ومكثَّف جداً، ولا يمكن للتطور التجريبي ولا الطبيعي أن يقترب من تحقيق عينات تسلسل بهذا العدد الهائل الذي يُحقِّقه التطوُّر الموجَّه، وبطبيعة الحال فإنَّ متواليات متغايرة للتطوُّر الطبيعي تقترب من متواليات البروتينات الوظيفيَّة وهذا ما يتمُّ تقليده في التطوُّر الموجَّه عن طريق تطفير جين فعَّال مسبقاً، يمكن تطفير الجين البدء انطلاقاً من نقطة معيَّنة عن طريق المُطفِّرات الكيميائيَّة أو التقنيَّات الأخرى[11] ، ويمكن محاكاة إعادة تجميع الجينات عن طريق خلط الحمض النووي باستخدام الإدخال والحذف بواسطة الترانسبوزونات[12]، لتحقيق منهج أكثر تركيزاً يعتمد على المعرفة الهيكليَّة والوظيفيَّة سوف تختلف مكتبة الجينات المُنتجة في نسبة المتغيرات الوظيفيَّة التي تحتوي عليها، وإذا ما استخدمنا كائناً حيَّاً للتعبير عن هذه الجينات فإنَّ التغيُّرات الموروثة في ذلك الجين ستبقي بقيَّة جينات الكائن الحي متماثلة بمعنى آخر فهي توفِّر بيئة وراثيَّة ثابتة ومستقرِّة.
الكشف عن الاختلافات
كما هو معروف تكون غالبيَّة الطفرات ضارَّة ولذلك فإنَّ مكتبات المُتغيِّرات تميل في الغالب إلى المتغيِّرات ذات النشاط المنخفض[13]، وهكذا سيكون اختبار الإنتاجيَّة العالية أمر حيوي لقياس النشاط الهادف للعثور على المتغيِّرات النادرة مع الطفرات المفيدة التي تحسِّن الخصائص المرغوب بها، توجد طريقتان رئيسيَّتان لعزل المتغيِّرات الوظيفيَّة: حيث تقوم أنظمة الانتقاء مباشرةً باختيار البروتينات الوظيفيَّة لبقاء الجين بينما تقوم أنظمة الفرز بفحص كل متغيِّر على حدة وتسمح بوضع حد كمِّي لفرز المتغيِّرات التي تؤدي للنشاط المرغوب، يمكن القيام بالانتقاء والفرز في الخلايا الحيَّة في الكائنات الحيَّة، أو بشكلٍ مباشر على البروتينات أو الرنا بدون الحاجة لوجود أي خلايا حتى في المختبر.[14][15]
خلال تطور الكائنات الحية يتمُّ تحويل بعض الجينات أو تغييرها ضمن الخليَّة الحيَّة مع الحفاظ على جميع الجينات الأخرى على حالها، يؤدي هذا التنسيق إلى اختيار خصائص معيَّنة ضمن البيئة الخلوية وهو أمرٌ مفيد عندما يستخدم البروتين المطوَّر أو الحمض النووي الريبي في الكائنات الحيَّة، أمَّا عندما يحدث التطوُّر الموجَّه في المختبر وليس في الكائن الحي فإنَّه يستخدم طريقة النسخ المختبرية لإنتاج البروتينات أو RNA في محلول صناعي مُعد خصيصاً لهذا الغرض، وبالطبع تكون هذه الطريقة أكثر فائدةً لأنَّها تحمل تنوُّعات أكثر وتحكُّم أكثر في الظروف التطوريَّة ويمكن القيام بالتجارب كذلك على البروتينات السامَّة للخلايا الحيَّة، بالإضافة لذلك يمكن لتجارب التطوُّر في المختبر أن تولِّد مكتبات تغيُّرات أكبر بكثير من تلك التي يمكن أن تحصل ضمن الخلايا الحيَّة في البيئة الطبيعيَّة والتي غالباً ما تكون محدودة.
الاختيار
تعتبر تقنية الاختيار بسيطة إلى حدٍّ ما حيث يُثبَّت الجزيء المستهدف على دعامة أو ركيزة معيَّنة حتى يتمُّ تدفُّق سلسلة من البروتينات المتباينة فوقه، فتعزل البروتينات غير المرغوب بها ويحتفظ بالتغيُّرات المتبقَّة لعزل جيناتها[16]، بواسطة هذا المنهج يتمُّ اختيار دورة واحدة من التحفيز من خلال جعل نشاط الأنزيم ضرورياً لبقاء الخلية على قيد الحياة إمَّا عن طريق تعديل مستقلبات حيوية أو تدمير مادة ضارة أو سامة، وهكذا يصبح نشاط الأنزيم هو عامل بقاء للخلية الحيَّة[17][18]، بالرغم من أنَّ أنظمة الاختيار هذه محدودة الإنتاجيَّة ولكنَّها أقل تكلفة، من جهة أخرى يصعب عادةً هندستها واستخلاص المعلومات عن الأنشطة الموجودة ضمن مكتبة المتغيِّرات بواسطتها.
التحري والكشف
نظام التحري والكشف والفرز هو بديل لطريقة الاختيار التي تمَّ شرحها سابقاً، في هذا النظام يتمُّ التعبير عن كلِّ جينة مختلفة بشكل فردي ومعايرتها لقياس النشاط كميَّاً ثمَّ يتمُّ ترتيب المتغيِّرات ويقرِّر المُجرِّب أي المتغيِّرات سيستخدمها كقوالب للدورة القادمة من دورات التطوُّر الموجَّه، وعلى الرغم من أنَّ معظم الفحوصات المستخدمة في نظام التحري والكشف تكون عادةً أقل تغطية من الناحية الكمية عند مقارنتها بطريقة الاختيار والعزل ولكنَّها تتمتَّع بميِّزات فريدة من نوعها لأنَّها تعطي معلومات تفصيلية عن كل متغيِّر من المتغيِّرات، ويمكن كذلك استخدام هذه البيانات المُصنَّفة والمفروزة لتمييز الأنشطة وتوزيعها وهو أمرٌ غير ممكن في أنظمة الاختيار البسيطة، وهكذا نجد أنَّ أنظمة الفرز لها مزايا هامَّة وخصوصاً عندما يتعلَّق الأمر بتوصيف تجريبي للتطوُّر التكيُّفي والتناسب مع البيئة المحيطة.
مزايا التطوُّر الموجَّه
يعتمد التخليق العقلاني للبروتين على معرفة مُسبقة متعمِّقة بهيكل البروتين وتركيبه بالإضافة إلى آليته التحفيزيَّة[17]، ثمَّ يتمُّ إجراء تغييرات مُحدَّدة عن طريق التطفير الموجَّه الهادف لتغيير وظيفة البروتين، مع الإشارة لوجود عيب هام في طريقة التطوُّر الموجَّه لأنَّه حتى عندما يكون هيكل وآلية عمل البروتين معروفين لا يزال من الصعب التنبؤ بالتغيير الناتج عن الطفرة، وربَّما تكون أهم ميزة للتطوُّر الموجَّه أنَّه لا توجد حاجة لفهم آلية النشاط المطلوب أو كيف تؤثر عليه الطفرات.[19]
حدود التطوُّر الموجَّه
هناك تقييد هام لحدود التطوُّر الموجَّه حيث يلزم إجراء اختبار عالي الإنتاجيَّة حتى نتمكَّن من قياس تأثيرات عدد كبير من الطفرات العشوائيَّة المختلفة، وهو الأمر الذي يتطلَّب القيام ببحث وتطوير مكثَّف قبل أن نتمكَّن من استخدام التطور الموجَّه، وغالباً ما تكون هذه القياسات خاضعة لمراقبة نشاط معيَّن وبالتالي فهي غير قابلة لتحويلها إلى تجار تطوُّر موجَّه جديدة.[20]
لفهم كيفيَّة تحقيق هذه التحسينات التطوريَّة يجب قياس خصائص الأنزيم المتطوِّر وتحسين نشاط المقايسة عن طريق زيادة نشاط التحفيز الأنزيمي أو زيادة تركيز الأنزيم المُحفِّز، بالإضافة لذلك لا يوجد ضمان أو تأكيد بأنَّ التحسين الذي يتمُّ إجراؤه على إحدى الركائز سيُحسِّن النشاط أو التحفيز المُجرى على الآخر، وتبدو أهميَّة ذلك عندما لا يمكن فحص النشاط المدروس أو تحفيزه بشكلٍ مباشر وعندها نستخدم الركيزة كبديل أو كمساعد لدراسة التجربة بدقة.[21]
تطبيقات عمليَّة
يستخدم التطوُّر الموجَّه على نطاقٍ واسع كبديل للتصميم العقلاني Rational design []، ويمكن أن يُستخدم أيضاً للإجابة عن الأسئلة الأساسيَّة المُتعلِّقة بتطوُّر الأنزيمات وتوضيحها بشكلٍ دقيق.[22]
في هندسة البروتينات
حقَّق التطور الموجَّه نجاحات هامَّة عند استخدامه كأداة لهندسة البروتينات وتعديلها، وكان الأفضل والأكثر نجاحاً في ثلاثة مجالات على وجه التحديد:
- تحسين الاستقرار الحيوي للبروتينات من أجل الاستخدام الحيوي التكنولوجي في درجات الحرارة المرتفعة أو عند استخدام مذيبات شديدة.[23][24]
- زيادة التقارب بين الأجسام العلاجيَّة المضادة [42]، وزيادة نشاط الأنزيمات المُصمَّمة.[25]
- تغيير خصائص الركيزة التي تربط الأنزيمات الموجودة للاستخدام في الصناعة غالباً.[26][27][28][29]
في أبحاث التطوُّر والدراسات المتعلقة به
تعتمد أبحاث التطور الطبيعي بشكلٍ تقليدي على الكائنات الحيَّة وجيناتها، ولكنَّ هذه الأبحاث تبقى محدودة بشكلٍ أساسي بسبب عدم وجود الحفريات التي تحتوي على سلاسل DNA القديمة، بالإضافة للمعرفة المنقوصة بالظروف البيئيَّة القديمة.
أمَّا التطوُّر الموجَّه فيدرس آليَّات التطوُّر ضمن نظام جيني خاضعة للرقابة الدقيقة على الأنزيمات المُفردة وطرق النسخ المتماثل[30][31]، على غرار التطوُّر التجريبي لحقيقيَّات النوى[32][33][34] ، وبدائيَّات النوى[35]، والفيروسات[36]، بالإضافة لذلك يسمح التطوُّر الموجَّه بالسيطرة على الضغوط التطوريَّة الانتخابيَّة، ومعدَّل الطفرات، وظروف البيئة المحيطة، وتشمل الظروف البيئة غير الحيَّة (مثل درجة الحرارة) والظروف البيئيَّة الحيَّة (الجينات الأخرى في الكائن الحي).
مراجع
- Lutz S (ديسمبر 2010)، "Beyond directed evolution--semi-rational protein engineering and design"، Current Opinion in Biotechnology، 21 (6): 734–43، doi:10.1016/j.copbio.2010.08.011، PMC 2982887، PMID 20869867.
- Cobb RE, Chao R, Zhao H (مايو 2013)، "Directed Evolution: Past, Present and Future"، AIChE Journal. American Institute of Chemical Engineers، 59 (5): 1432–1440، doi:10.1002/aic.13995، PMC 4344831، PMID 25733775.
- Mills DR, Peterson RL, Spiegelman S (يوليو 1967)، "An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule"، Proceedings of the National Academy of Sciences of the United States of America، 58 (1): 217–24، doi:10.1073/pnas.58.1.217، PMC 335620، PMID 5231602.
- Hall BG (يوليو 1978)، "Experimental evolution of a new enzymatic function. II. Evolution of multiple functions for ebg enzyme in E. coli"، Genetics، 89 (3): 453–65، PMC 1213848، PMID 97169.
- Smith GP (يونيو 1985)، "Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface"، Science، 228 (4705): 1315–7، doi:10.1126/science.4001944، PMID 4001944.
- Chen, Keqin؛ Arnold, Frances H. (1991)، "Enzyme Engineering for Nonaqueous Solvents: Random Mutagenesis to Enhance Activity of Subtilisin E in Polar Organic Media"، Bio/Technology (باللغة الإنجليزية)، 9 (11): 1073–1077، doi:10.1038/nbt1191-1073، ISSN 0733-222X.
- Kim, Eun-Sung (27 نوفمبر 2008)، "Directed Evolution: A Historical Exploration into an Evolutionary Experimental System of Nanobiotechnology, 1965–2006"، Minerva (باللغة الإنجليزية)، 46 (4): 463–484، doi:10.1007/s11024-008-9108-9، ISSN 0026-4695.
- Packer MS, Liu DR (يوليو 2015)، "Methods for the directed evolution of proteins"، Nature Reviews. Genetics (باللغة الإنجليزية)، 16 (7): 379–94، doi:10.1038/nrg3927، PMID 26055155.
- "The Nobel Prize in Chemistry 2018"، NobelPrize.org (باللغة الإنجليزية)، مؤرشف من الأصل في 28 مارس 2019، اطلع عليه بتاريخ 03 أكتوبر 2018.
- Voigt CA, Kauffman S, Wang ZG (2000)، Rational evolutionary design: the theory of in vitro protein evolution، Advances in Protein Chemistry، ج. 55، ص. 79–160، doi:10.1016/s0065-3233(01)55003-2، ISBN 9780120342556، PMID 11050933.
- Lipovsek D, Plückthun A (يوليو 2004)، "In-vitro protein evolution by ribosome display and mRNA display"، Journal of Immunological Methods، 290 (1–2): 51–67، doi:10.1016/j.jim.2004.04.008، PMID 15261571.
- Dryden DT, Thomson AR, White JH (أغسطس 2008)، "How much of protein sequence space has been explored by life on Earth?"، Journal of the Royal Society, Interface، 5 (25): 953–6، doi:10.1098/rsif.2008.0085، PMC 2459213، PMID 18426772.
- Hartl DL (أكتوبر 2014)، "What can we learn from fitness landscapes?"، Current Opinion in Microbiology، 21: 51–7، doi:10.1016/j.mib.2014.08.001، PMC 4254422، PMID 25444121.
- Badran AH, Liu DR (فبراير 2015)، "In vivo continuous directed evolution"، Current Opinion in Chemical Biology، 24: 1–10، doi:10.1016/j.cbpa.2014.09.040، PMC 4308500، PMID 25461718.
- Kumar A, Singh S (ديسمبر 2013)، "Directed evolution: tailoring biocatalysts for industrial applications"، Critical Reviews in Biotechnology، 33 (4): 365–78، doi:10.3109/07388551.2012.716810، PMID 22985113.
- Willats WG (ديسمبر 2002)، "Phage display: practicalities and prospects"، Plant Molecular Biology، 50 (6): 837–54، doi:10.1023/A:1021215516430، PMID 12516857.
- Leemhuis H, Stein V, Griffiths AD, Hollfelder F (أغسطس 2005)، "New genotype-phenotype linkages for directed evolution of functional proteins"، Current Opinion in Structural Biology، 15 (4): 472–8، doi:10.1016/j.sbi.2005.07.006، PMID 16043338.
- Verhoeven KD, Altstadt OC, Savinov SN (مارس 2012)، "Intracellular detection and evolution of site-specific proteases using a genetic selection system"، Applied Biochemistry and Biotechnology، 166 (5): 1340–54، doi:10.1007/s12010-011-9522-6، PMID 22270548.
- Nguyen AW, Daugherty PS (مارس 2005)، "Evolutionary optimization of fluorescent proteins for intracellular FRET"، Nature Biotechnology، 23 (3): 355–60، doi:10.1038/nbt1066، PMID 15696158.
- Schaerli Y, Hollfelder F (ديسمبر 2009)، "The potential of microfluidic water-in-oil droplets in experimental biology"، Molecular BioSystems، 5 (12): 1392–404، doi:10.1039/b907578j، PMID 20023716.
- Marshall SA, Lazar GA, Chirino AJ, Desjarlais JR (مارس 2003)، "Rational design and engineering of therapeutic proteins"، Drug Discovery Today، 8 (5): 212–21، doi:10.1016/s1359-6446(03)02610-2، PMID 12634013.
- Wilson CJ (27 أكتوبر 2014)، "Rational protein design: developing next-generation biological therapeutics and nanobiotechnological tools"، Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology، 7 (3): 330–41، doi:10.1002/wnan.1310، PMID 25348497.
- Gatti-Lafranconi P, Natalello A, Rehm S, Doglia SM, Pleiss J, Lotti M (يناير 2010)، "Evolution of stability in a cold-active enzyme elicits specificity relaxation and highlights substrate-related effects on temperature adaptation"، Journal of Molecular Biology، 395 (1): 155–66، doi:10.1016/j.jmb.2009.10.026، PMID 19850050.
- Zhao H, Arnold FH (يناير 1999)، "Directed evolution converts subtilisin E into a functional equivalent of thermitase"، Protein Engineering، 12 (1): 47–53، doi:10.1093/protein/12.1.47، PMID 10065710.
- Hawkins RE, Russell SJ, Winter G (أغسطس 1992)، "Selection of phage antibodies by binding affinity. Mimicking affinity maturation"، Journal of Molecular Biology، 226 (3): 889–96، doi:10.1016/0022-2836(92)90639-2، PMID 1507232.
- Shaikh FA, Withers SG (أبريل 2008)، "Teaching old enzymes new tricks: engineering and evolution of glycosidases and glycosyl transferases for improved glycoside synthesis"، Biochemistry and Cell Biology، 86 (2): 169–77، doi:10.1139/o07-149، PMID 18443630.
- Cheriyan M, Walters MJ, Kang BD, Anzaldi LL, Toone EJ, Fierke CA (نوفمبر 2011)، "Directed evolution of a pyruvate aldolase to recognize a long chain acyl substrate"، Bioorganic & Medicinal Chemistry، 19 (21): 6447–53، doi:10.1016/j.bmc.2011.08.056، PMC 3209416، PMID 21944547.
- MacBeath G, Kast P, Hilvert D (مارس 1998)، "Redesigning enzyme topology by directed evolution"، Science، 279 (5358): 1958–61، doi:10.1126/science.279.5358.1958، PMID 9506949.
- Toscano MD, Woycechowsky KJ, Hilvert D (2007)، "Minimalist active-site redesign: teaching old enzymes new tricks"، Angewandte Chemie، 46 (18): 3212–36، doi:10.1002/anie.200604205، PMID 17450624.
- Pääbo S, Poinar H, Serre D, Jaenicke-Despres V, Hebler J, Rohland N, Kuch M, Krause J, Vigilant L, Hofreiter M (2004)، "Genetic analyses from ancient DNA"، Annual Review of Genetics، 38 (1): 645–79، doi:10.1146/annurev.genet.37.110801.143214، PMID 15568989.
- Höss M, Jaruga P, Zastawny TH, Dizdaroglu M, Pääbo S (أبريل 1996)، "DNA damage and DNA sequence retrieval from ancient tissues"، Nucleic Acids Research، 24 (7): 1304–7، doi:10.1093/nar/24.7.1304، PMC 145783، PMID 8614634.
- Bloom JD, Arnold FH (يونيو 2009)، "In the light of directed evolution: pathways of adaptive protein evolution"، Proceedings of the National Academy of Sciences of the United States of America، 106 Suppl 1 (Supplement_1): 9995–10000، doi:10.1073/pnas.0901522106، PMC 2702793، PMID 19528653.
- Moses AM, Davidson AR (مايو 2011)، "In vitro evolution goes deep"، Proceedings of the National Academy of Sciences of the United States of America، 108 (20): 8071–2، doi:10.1073/pnas.1104843108، PMC 3100951، PMID 21551096.
- Goldsmith M, Tawfik DS (أغسطس 2012)، "Directed enzyme evolution: beyond the low-hanging fruit"، Current Opinion in Structural Biology، 22 (4): 406–12، doi:10.1016/j.sbi.2012.03.010، PMID 22579412.
- Salehi-Ashtiani K, Szostak JW (نوفمبر 2001)، "In vitro evolution suggests multiple origins for the hammerhead ribozyme"، Nature، 414 (6859): 82–4، doi:10.1038/35102081، PMID 11689947.
- Sumper M, Luce R (يناير 1975)، "Evidence for de novo production of self-replicating and environmentally adapted RNA structures by bacteriophage Qbeta replicase"، Proceedings of the National Academy of Sciences of the United States of America، 72 (1): 162–6، doi:10.1073/pnas.72.1.162، PMC 432262، PMID 1054493.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري