تعدد الصيغ الصبغية
تعدد الصيغ الصبغية (بالإنجليزية: polyploidy) هي حالة تتميز باحتواء خلية الكائن الحي على أكثر من مجموعتين (متماثلتين) من الصبغيات. تعد غالبية الأنواع التي تمتلك نوى (حقيقيات النوى) ثنائية الصيغة الصبغية، أي أنها تحوي مجموعتين من الصبغيات، تُورث كل مجموعة من أحد الأبوين. لكن، تكون بعض الكائنات العضوية متعددة الصيغ الصبغية، ويعتبر هذا الأمر شائعًا في النباتات على وجه الخصوص. تتمتع معظم وحيدات النوى بخلايا جسمية ذات صيغة صبغية ثنائية ولكنها تنتج جاميت/أمشاج (بويضات ونطاف) أحادية الصيغة الصبغية عبر عملية الانقسام المنصف. تحوي أحاديات الصيغة الصبغية على مجموعة واحدة فقط من الصبغيات، وينطبق المصطلح على الخلايا أو الكائنات الحية التي تحوي نصف العدد الصبغي عادةً. يعد ذكور النحل وأنواع أخرى من غشائيات الأجنحة على سبيل المثال من أحاديات الصيغة الصبغية. على عكس الحيوانات، تمتلك النباتات والطحالب عديدة الخلايا دورات حياة فيها جيلان عديدا الخلايا متناوبان. يكون جيل الطور المشيجي أحادي الصيغة الصبغية، وينتج الأمشاج عبر الانقسام الخيطي، أما الجيل البوغي يكون ثنائي الصيغة الصبغية وينتج الأبواغ عبر الانقسام المنصف.
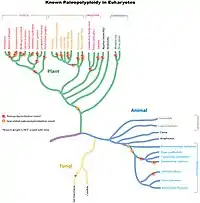
قد يحدث تعدد الصيغ الصبغية نتيجة انقسام خلوي شاذ، إما خلال الانقسام الخيطي أو في الطور الأول من الانقسام المنصف عادةً (قد ينشأ من فشل الصبغيات في الانفصال خلال الانقسام المنصف أو من تلقيح البويضة بأكثر من نطفة[1]). بالإضافة إلى ذلك، يمكن تحريض تعدد الصيغ الصبغية في النباتات والمزارع الخلوية بواسطة بعض المواد الكيميائية، وأشهرها الكولشيسين، الذي يؤدي إلى تضاعف الصبغيات على الرغم من أن استخدامه يسفر عن عدة نتائج أخرى أقل وضوحًا. يؤدي الأوريزالين أيضًا إلى تضاعف المحتوى الصبغي الموجود.
يحدث تعدد الصيغ الصبغية في الأنسجة البشرية المتمايزة في الكبد وعضلة القلب ونقي العظم والمشيمة.[2] يظهر في الخلايا الجسمية لبعض الحيوانات مثل السمك الذهبي[3] والسلمون والسلمندر، ولكنه يعد أكثر شيوعًا في السراخس والنباتات المزهرة (خطمي وردي صيني)، بما في ذلك النوعان البري والمزروع. خضع القمح على سبيل المثال للتهجين والتعديل على يد البشر على امتداد آلاف السنين، وبات يأتي بسلالات ثنائية الصيغة الصبغية (مجموعتان من الصبغيات) ورباعية الصيغة الصبغية (أربع مجموعات من الصبغيات) وتُعرف هذه السلالة بالقمح الصلب أو قمح المعكرونة، وسلالة سداسية الصيغة الصبغية (ست مجموعات من الصبغيات) وتُعرف باسم قمح الخبز. تعتبر عدة أنواع هامة زراعيًا من جنس براسيكا أيضًا رباعية الصيغة الصبغية.
قد يمثل تعدد الصيغ الصبغية آلية للانتواع المستوطن لأن الأنواع متعددة الصيغة الصبغية غير قابلة للتهجين مع أسلافها ثنائية الصيغة الصبغية. يعد نبات الزهرة المقنعة الكاملة مثالًا على ذلك. أكد التسلسل أن هذا النوع نشأ من المقنعة الروبرتسي، وهو جنس نقي ثلاثي الصيغة الصبغية بين المقنعة المنقطة والمقنعة الصفراء وهما نوعان طُرحا وجُنّسا في المملكة المتحدة. ظهرت مجموعات جديدة من المقنعة الكاملة في الأراضي الوسطى في اسكتلندا وجزر أوركني عبر التضاعف الجينومي في المجموعات المحلية للمقنعة الروبرتسي.[4] تعد المقنعة الكاملة غير نقية بسبب الطفرات الجينية النادرة.[5]
المصطلحات
الأنواع
تُصنف الأنواع متعددة الصيغ الصبغية وفقًا لعدد مجموعات الصبغيات في النواة. يُستخدم الحرف إكس لتمثيل عدد الصبغيات في المجموعة الواحدة:
- أحادية الصيغة الصبغية (مجموعة واحدة، ×1).
- ثنائية الصيغة الصبغية (مجموعتان، ×2).
- ثلاثية الصيغة الصبغية (ثلاث مجموعات، ×3)، مثل الزعفران المزروع والبطيخ عديم البذر، وتعد شائعة أيضًا في شعبة بطيئات الخطو.[6]
- رباعية الصيغة الصبغية (أربع مجموعات، ×4)، مثل سمك السلمونيات[7] والقطن الزغبي.[8]
- خماسية الصيغة الصبغية (خمس مجموعات، ×5)، مثل البيرش كيناي (البتولا كيناي).
- سداسية الصيغة الصبغية (ست مجموعات، ×6)، مثل القمح والكيوي.[9]
- سباعية الصيغة الصبغية (سبع مجموعات، ×7).
- ثمانية الصيغة الصبغية (ثمان مجموعات، ×8)، مثل جنس الحفش (جنس من فصيلة الحفشيات) والأضاليات.
- عشارية الصيغة الصبغية (عشر مجموعات، ×10)، مثل بعض أنواع الفراولة.
- اثنا عشر صيغة صبغية (اثنا عشر مجموعة، ×12)، مثل نباتي عرف الديك وعشب الحبل الشائع أو القيطم الأوغندي البرمائي.[10]
التعدد الصبغي الذاتي
تعتبر الكائنات ذات التعدد الصبغي الذاتي عديدات صيغة صبغية تمتلك عدة مجموعات صبغية مشتقة من أصنوفة واحدة.
تضم الأمثلة على التعدد الصبغي الذاتي الطبيعي النبات الطفيلي المعروف باسم التوليميا المنزية[11] والحفش الأبيض المعروف باسم أسيبنسر ترانسموناتوم.[12] تنتج معظم حالات التعدد الصبغي الذاتي عن اندماج الأمشاج غير المختزلة (2ن)، ما يؤدي إلى نسل ثلاثي الصيغة (ن + 2ن = 3ن) أو نسل رباعي الصيغة الصبغية (2ن + 2ن = 4ن).[13] يكون النسل ثلاثي الصيغة عقيمًا عادةً (مثل ظاهرة «العائق الثلاثي»)، ولكن قد ينتج نسبًا عالية من الأمشاج غير المختزلة في بعض الحالات وبالتالي يساعد في تكوين صيغ رباعية الصبغيات. يشار إلى الطريق الذي يعطي رباعيات الصيغة الصبغية باسم «الجسر ثلاثي الصيغة الصبغية». قد تستمر ثلاثيات الصيغة الصبغية أيضًا من خلال التكاثر اللاجنسي.[13] في الواقع، غالبًا ما يرتبط التثلث الصبغي الذاتي المستقر في النباتات بأنظمة التزاوج الوراثي. في النظم الزراعية،[14] قد يؤدي التثلث الصبغي الذاتي إلى غياب البذور، كما هو الحال في البطيخ والموز.[15] يُستخدم تثلث الصبغيات في استزراع السلمون والسلمون المرقط للحث على العقم.[16][17]
نادرًا ما يحدث التعدد الصبغي الذاتي من تضاعف الجينوم الجسمي العفوي، والذي لوحظ في شذوذات برعم التفاح (التفاح المنزلي).[18] يعد هذا المسار أيضًا المسار الأشهر لتعدد الصيغ الصبغية المحرض صناعيًا، إذ تُستخدم طرق مثل اندماج البروتوبلاست أو العلاج بالكولشيسين أو الأوريزالين أو مثبطات الانقسام الخيطي لتعطيل الانقسام الخيطي الطبيعي، ما يؤدي إلى إنتاج الخلايا متعددة الصيغ الصبغية. يمكن أن تكون هذه العملية مفيدة في تكاثر النبات، خاصة عند محاولة إدخال الأصول الوراثية عبر المستويات التضاعفية.[19]
تمتلك عديدات الصيغ الصبغية ما لا يقل عن ثلاث مجموعات صبغية متماثلة، والتي قد تؤدي إلى معدلات عالية من الاقتران متعدد التكافؤ أثناء الانقسام المنصف (خاصة في عديدات الصيغة الصبغية الذاتية المتكونة حديثًا، والمعروفة أيضًا باسم عديدات الصيغة الصبغية الحديثة) وإلى انخفاض مرافق في الخصوبة بسبب إنتاج الأمشاج مختلة الصيغة الصبغية.[20] يمكن أن يؤدي الانتقاء الطبيعي أو الاصطناعي للخصوبة إلى استقرار سريع للانقسام المنصف في عديدات الصيغة الصبغية الذاتية عن طريق استعادة الاقتران ثنائي التكافؤ أثناء الانقسام المنصف، لكن تدفع الدرجة العالية من التماثل بين الصبغيات المتضاعفة بعديدات الصيغة الصبغية إلى إظهار الموروث متعدد الصبغيات.[21] تُستخدم هذه السمة أغلب الأحيان بمثابة معيار تشخيصي لتمييز عديدات الصيغة الصبغية الذاتية عن عديدات الصيغة الصبغية المتغايرة، والتي تظهر عادةً موروثًا ثنائي الصبغي عقب تقدمها بعد مرحلة عديدات الصيغة الصبغية الحديثة.[22] رغم أن معظم الأنواع متعددة الصيغ الصبغية توصف وصفًا لا لبس فيه فتكون إما متعددة صيغة صبغية ذاتية أو متعددة صيغة صبغية مغايرة، تمثل هاتان الفئتان طرفي طيف الاختلاف بين الجينومات الفرعية الأبوية. قد تعرض عديدات الصيغ الصبغية الواقعة بين هذين الطرفين، والتي يشار إليها غالبًا باسم عديدات الصيغ الصبغية المتغايرة القطاعية، مستويات وسيطة من الموروث متعدد الصبغيات والتي قد تختلف حسب الموقع.[23][24]
يُعتقد أن حوالي نصف العدد الكلي لعديدات الصيغة الصبغية ناتج عن تعدد صيغة صبغية ذاتي،[25][26] على الرغم من أن العديد من العوامل تجعل تقدير هذه النسبة صعبًا.[27]
انظر أيضًا
مراجع
- Solomon, Eldra (2014)، Solomon/Martin/Martin/Berg, BIOLOGY، Cengage Learning، ص. 344، ISBN 978-1285423586.
- Velicky, P.؛ Meinhardt, G.؛ Plessl, K.؛ Vondra, S.؛ Weiss, T.؛ Haslinger, P.؛ Lendl, T.؛ Aumayr, K.؛ Mairhofer, M.؛ Zhu, X.؛ Schütz, B.؛ Hannibal, R. L.؛ Lindau, R.؛ Weil, B.؛ Ernerudh, J.؛ Neesen, J.؛ Egger, G.؛ Mikula, M.؛ Röhrl, C.؛ Urban, A. E.؛ Baker, J.؛ Knöfler, M.؛ Pollheimer, J. (2018)، "Genome amplification and cellular senescence are hallmarks of human placenta development"، PLOS Genetics، 14 (10): e1007698، doi:10.1371/journal.pgen.1007698، PMC 6200260، PMID 30312291.
- Ohno, S.؛ Muramoto, J.؛ Christian, L.؛ Atkin, N. B. (1967)، "Diploid-tetraploid relationship among old-world members of the fish family Cyprinidae"، Chromosoma، 23 (1): 1–9، doi:10.1007/BF00293307، S2CID 1181521.
- Vallejo Marín, M.؛ Buggs, R. J. A.؛ Cooley, A. M.؛ Puzey, J. R. (2015)، "Speciation by genome duplication: Repeated origins and genomic composition of the recently formed allopolyploid species Mimulus peregrinus"، Evolution، 69 (6): 1487–1500، doi:10.1111/evo.12678، PMC 5033005، PMID 25929999.
- Fessenden, M.، "Make Room for a New Bloom: New Flower Discovered"، Scientific American، مؤرشف من الأصل في 9 أكتوبر 2021، اطلع عليه بتاريخ 22 فبراير 2017.
- Bertolani, R. (2001)، "Evolution of the reproductive mechanisms in Tardigrades: a review"، Zoologischer Anzeiger، 240 (3–4): 247–252، doi:10.1078/0044-5231-00032.
- Stouder, D. J.؛ Bisson, P. A.؛ Naiman, R. J. (1997)، Pacific Salmon and Their Ecosystems: Status and Future Options، Springer، ص. 30–31، ISBN 978-0-412-98691-8، مؤرشف من الأصل في 4 سبتمبر 2021، اطلع عليه بتاريخ 09 يوليو 2013.
- Adams, K. L.؛ Wendel, J. F. (2005)، "Polyploidy and genome evolution in plants"، Current Opinion in Plant Biology، 8 (2): 135–141، doi:10.1016/j.pbi.2005.01.001، PMID 15752992.
- Crowhurst, R. N.؛ Whittaker, D.؛ Gardner, R. C.، "The genetic origin of kiwifruit"، مؤرشف من الأصل في 25 فبراير 2021.
- Ainouche, M. L.؛ Fortune, P. M.؛ Salmon, A.؛ Parisod, C.؛ Grandbastien, M.-A.؛ Fukunaga, K.؛ Ricou, M.؛ Misset, M.-T. (2008)، "Hybridization, polyploidy and invasion: Lessons from Spartina (Poaceae)"، Biological Invasions، 11 (5): 1159–1173، doi:10.1007/s10530-008-9383-2، S2CID 25522023.
- Soltis, D. E. (01 يناير 1984)، "Autopolyploidy in Tolmiea menziesii (Saxifragaceae)"، American Journal of Botany، 71 (9): 1171–1174، doi:10.2307/2443640، JSTOR 2443640.
- Drauch Schreier, A.؛ Gille, D.؛ Mahardja, B.؛ May, B. (01 نوفمبر 2011)، "Neutral markers confirm the octoploid origin and reveal spontaneous autopolyploidy in white sturgeon, Acipenser transmontanus"، Journal of Applied Ichthyology (باللغة الإنجليزية)، 27: 24–33، doi:10.1111/j.1439-0426.2011.01873.x، ISSN 1439-0426.
- Bretagnolle, F.؛ Thompson, J. D. (01 يناير 1995)، "Gametes with the somatic chromosome number: mechanisms of their formation and role in the evolution of autopolyploid plants"، New Phytologist (باللغة الإنجليزية)، 129 (1): 1–22، doi:10.1111/j.1469-8137.1995.tb03005.x، ISSN 1469-8137، PMID 33874422.
- Müntzing, Arne (مارس 1936)، "The Evolutionary Significance of Autopolyploidy"، Hereditas (باللغة الإنجليزية)، 21 (2–3): 363–378، doi:10.1111/j.1601-5223.1936.tb03204.x، ISSN 1601-5223.
- Varoquaux, F.؛ Blanvillain, R.؛ Delseny, M.؛ Gallois, P. (يونيو 2000)، "Less is better: new approaches for seedless fruit production"، Trends in Biotechnology، 18 (6): 233–242، doi:10.1016/s0167-7799(00)01448-7، ISSN 0167-7799، PMID 10802558.
- Lincoln, R. F.؛ Scott, A. P. (1983)، "Production of all-female triploid rainbow trout"، Aquaculture (باللغة الإنجليزية)، 30 (1–4): 375–380، doi:10.1016/0044-8486(83)90179-5.
- Cotter, D.؛ O'Donovan, V.؛ Ó Maoiléidigh, N.؛ Rogan, G.؛ Roche, N.؛ Wilkins, N. P. (يونيو 2000)، "An evaluation of the use of triploid Atlantic salmon (Salmo salar L.) in minimising the impact of escaped farmed salmon on wild populations"، Aquaculture، 186 (1–2): 61–75، doi:10.1016/S0044-8486(99)00367-1.
- Dermen, H. (مايو 1951)، "Tetraploid and Diploid Adventitious Shoots: From a Giant Sport of McIntosh Apple"، Journal of Heredity، 42 (3): 145–149، doi:10.1093/oxfordjournals.jhered.a106189، ISSN 0022-1503.
- Dwivedi, S. L.؛ Upadhyaya, H. D.؛ Stalker, H. T.؛ Blair, M. W.؛ Bertioli, D. J.؛ Nielen, S.؛ Ortiz, R. (يناير 2007)، Janick, Jules (المحرر)، Plant Breeding Reviews (PDF) (باللغة الإنجليزية)، John Wiley & Sons، ص. 179–230، doi:10.1002/9780470380130.ch3، ISBN 9780470380130، مؤرشف من الأصل (PDF) في 24 فبراير 2021.
- Justin, R. (يناير 2002)، "Neopolyploidy in Flowering Plants"، Annual Review of Ecology and Systematics، 33 (1): 589–639، doi:10.1146/annurev.ecolsys.33.010802.150437، S2CID 41689384، مؤرشف من الأصل في 3 سبتمبر 2021.
- Parisod, C.؛ Holderegger, R.؛ Brochmann, C. (أبريل 2010)، "Evolutionary consequences of autopolyploidy"، The New Phytologist، 186 (1): 5–17، doi:10.1111/j.1469-8137.2009.03142.x، ISSN 1469-8137، PMID 20070540.
- Le Comber, S. C.؛ Ainouche, M. L.؛ Kovarik, A.؛ Leitch, A. R. (أبريل 2010)، "Making a functional diploid: from polysomic to disomic inheritance"، The New Phytologist، 186 (1): 113–122، doi:10.1111/j.1469-8137.2009.03117.x، ISSN 1469-8137، PMID 20028473.
- Stebbins, G. L. (1950)، Variation and Evolution in Plants، Oxford University Press.[بحاجة لرقم الصفحة]
- Stebbins, G. L. (1947)، Types of Polyploids: Their Classification and Significance، Advances in Genetics (باللغة الإنجليزية)، ج. 1، ص. 403–429، doi:10.1016/s0065-2660(08)60490-3، ISBN 9780120176014، PMID 20259289.
- Ramsey, J.؛ Schemske, D. W. (01 يناير 1998)، "Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants"، Annual Review of Ecology and Systematics، 29 (1): 467–501، doi:10.1146/annurev.ecolsys.29.1.467، S2CID 31637733، مؤرشف من الأصل في 3 سبتمبر 2021.
- Barker, M. S.؛ Arrigo, N.؛ Baniaga, A. E.؛ Li, Z.؛ Levin, D. A. (01 أبريل 2016)، "On the relative abundance of autopolyploids and allopolyploids"، New Phytologist (باللغة الإنجليزية)، 210 (2): 391–398، doi:10.1111/nph.13698، ISSN 1469-8137، PMID 26439879.
- Doyle, J. J.؛ Sherman-Broyles, S. (01 يناير 2017)، "Double trouble: taxonomy and definitions of polyploidy"، New Phytologist (باللغة الإنجليزية)، 213 (2): 487–493، doi:10.1111/nph.14276، ISSN 1469-8137، PMID 28000935.
 بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء