جهد الراحة
عندما يكون جهد الغشاء ثابتًا، نسبيًا، وتكون الخليّة هامدة (أي في حالة غير فعّآلة كهربائيًا)، ينعت هذا الجهد بجهد الراحة للغشاء، وهذا بعكس حالات أخرى ككمون الفعل أو التغيير التدريجي في الجهد الغشائي والتي هي بمثابة ظواهر كهركيميائية ديناميّة.[1][2][3] من ناحية فيزيائية-حيوية، ليس هنالك أي فرق بين جهد الراحة والظواهر الديناميّة ككمون الفعل، فكل هذه الظواهر سببها الأساسي هو التغييرات في نفاذية غشاء الخلية لأيونات البوتاسيوم والصوديوم والكالسيوم والكلور، والتي تحدث جراء التغييرات المتناسقة في القدرات الوظيفية للقنوات الشاردية والمضخات الشاردية وناقلات وحاملات الأيونات. إذا وجدت عدة أنواع من الأيونات في الوسط الخارجي للعصبون وفي الهيولى، فيكون لكل منها جهد اعتكاس الذي يتحدد وفق معادلة نرنست. لكنّ جهد الراحة للغشاء، أي الجهد الذي لا يوجد فيه أي تيّار أيونات صافٍ عبر الغشاء يختلف عادة عن أي من جهود الاعتكاس. عوضًا عن ذلك، فإنّ جهد الراحة يتحدد وفق معادلة غولدمان التي تأخذ بالحسبان كل أنواع وتراكيز الأيونات الموجودة، بالإضافة إلى النفاذية النسبية لكل أيون عبر الغشاء الهيولي. وإنّ معادلة غولدمان بالنسبة لثلاثة الأيونات وحيدة القيمة (صوديوم وبوتاسيوم وكالسيوم) الأهم بالنسبة لجهد الغشاء في كمون الفعل:

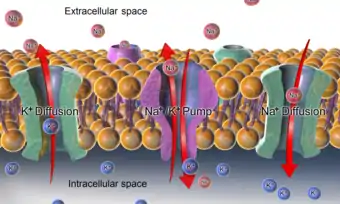
حيث الإشارات الرياضية مطابقة للإشارات في القسم السابق، وPi هي النفاذية النسبية للأيون i عبر الغشاء الهيولي. يجدر الذكر أنّ بسبب كون أيون الكلور ذا شحنة سالبة، فإنّ تأثيره في المعادلة معاكس لتأثير الأيونات الأخرى؛ تركيزه خارج الخلية يظهر في المقام، في حين يظهر تركيزه داخل الخلية في البسط. بالإضافة، فإذا أردنا إدراج تأثير أيون الكالسيوم في المعادلة، وهو تأثير بالغ الأهمية إذا ما نظرنا إلى كمون الفعل في خلية عضلية، فإنّ معادلة جهد التوازن تصبح أكثر معقّدة. بالإضافة، لغالبية الخلايا الحيوانية الهامدة، فإنّ الغشاء الهيولي نَفوذ لأيون البوتاسيوم أكثر بكثير من أي من الأيونات الأخرى المهمة، ولذا يكون جهد الراحة أقرب إلى جهد الاعتكاس لأيون البوتاسيوم منه إلى الأيونات الأخرى. لتوضيح النقطة السابقة، لنفرض أنّ خلية ما موجودة في وسط يحتوي على أيونات بوتاسيوم وصوديوم فحسب، وأنّ السائل الهيولي الداخلي للخلية يحتوي على هذين الأيونين فقط هو الآخر. بالإضافة إلى ذلك، لنفرض أنّ للأيونات نفاذية نسبية متساوية، وأنّ تراكيزهما متساوية لكنها معكوسة (أي تركيز البوتاسيوم داخل الخلية يساوي تركيز الصوديوم خارج الخلية والعكس أيضًا)، بما معناه أنّ التدفق الصافي لكل من الأيونات عبر غشاء الخلية متساوي فيما عدا إتجاه هذا التدفق. في هذه الحالة، سيكون لأيونات البوتاسيوم جهد اعتكاس معيّن، E_K، في حين يكون لأيونات الصوديوم جهد اعتكاس يحقّق: E_{Na} = - E_k. بسبب التماثل بين الأيونين في هذه الحالة، فإنّ جهد الراحة للخلية سيساوي المتوسط الحسابي لجهدي اعتكاس الأيونين، أي صفر في هذه الحالة. طبعًا، فبالإمكان استخلاص نفس النتيجة من معادلة غولدمان أعلاه. مع هذا، فيجب التنويه إلى أنّ هذه حالة من التوازن غير المستقر، حيث أنّ جهد يعادل صفرًا لا يؤدي إلى عدم وجود تيارات أيونية عبر غشاء الخلية، بل العكس هو الصحيح، إذ أنّ الأيونات ستستمر في التدفق وفق تدرج تراكيزها، ولكن جهد الغشاء يبقى ثابتًا بفعل مضخات شاردية تعمل بعكس تدرج التراكيز لأيونات البوتاسيوم والصوديوم، فتعمل على إدخال أيونات البوتاسيوم إلى الخلية وإخراج أيونات الصوديوم منها. عمل هذه المضخات يحتاج إلى جزيئات أدينوزين ثلاثي الفوسفات، ولهذا السبب فإنّ مثل الحالة الموصوفة أعلاه والتي تتساوي فيها التفاذية النسبية للأيونات هي ذات فعالية منخفضة. لذا، ففي الخلية الحيوية تكون هنالك نفاذية نسبية عالية لأيون البوتاسيوم (أي أن جهد الراحة يكون قريب لجهد اعتكاس البوتاسيوم)، في حين يتم التلاعب بجهد الغشاء بواسطة تغيير النفاذية النسبية للأيونات الأخرى، كالصوديوم أو الكلور (مثلما يحدث في كمون الفعل)، عن طريق فتح أو إغلاق القنوات الشاردية، على سبيل المثال. في الوضع الفيزيولوجي لخلية حيوانيّة تعادل نفاذية أيون الصوديوم حوالي %5 من نفاذية أيون البوتاسيوم. هذا الأمر يجعل جهد الراحة للخلية قريبًا من جهد اعتكاس أيون البوتاسيوم مع ميل بسيط نحو الصفر، أي -73 \ mV. نقطة مهمّة أخرى هي كون الطبقة الثنائية الدسمة ذات نفاذية صغيرة جدًا للماء ولأيونات البوتاسيوم والصوديوم. مع ذلك، فإنّ الغشاء يحتوي على العديد من قنوات المياه والقنوات والمضخات الأيونية ومبادلات الأيونات وناقلات الأيونات والتي تمكن من زيادة النفاذية النسبية لبعض الأيونات أو لجزيئات الماء.
مراجع
- "Muscles"، users.rcn.com، 24 يناير 2015، مؤرشف من الأصل في 07 نوفمبر 2015، اطلع عليه بتاريخ 01 يونيو 2016.
- exampleof an فيزيولوجيا كهربائية experiment to demonstrate the importance of K+ for the resting potential. The dependence of the resting potential on the extracellular concentration of K+ is shown in Figure 2.6 of Neuroscience, 2nd edition, by Dale Purves, George J. Augustine, David Fitzpatrick, Lawrence C. Katz, Anthony-Samuel LaMantia, James O. McNamara, S. Mark Williams. Sunderland (MA): Sinauer Associates, Inc.; 2001. نسخة محفوظة 11 يناير 2020 على موقع واي باك مشين.
- Seyfarth, Ernst-August (01 يناير 2006)، "Julius Bernstein (1839-1917): pioneer neurobiologist and biophysicist"، Biological Cybernetics، 94 (1): 2–8، doi:10.1007/s00422-005-0031-y، ISSN 0340-1200، PMID 16341542.
 بوابة طب
بوابة طب بوابة علم الأحياء
بوابة علم الأحياء بوابة علوم عصبية
بوابة علوم عصبية