خلية حارسة
الخلايا الحارسة (بالإنجليزية: Guard cells) الخلايا الحارسة هي خلايا نباتية متخصصة موجودة في بشرة الأوراق والسيقان وغيرها من الأعضاء التي تُستخدم للتحكم بالتبادل الغازي. تُنتج هذه الخلايا في أزواج يوجد بين كل خليتين فيها فجوة تسمى الثغر أو السُم. تتسع هذه المسام عندما يكون الماء متاحًا بوفرة إذ تكون الخلايا الحارسة منتفخة، وتغلق عندما يقل الماء كثيرًا إذ تكون الخلايا الحارسة منكمشة. يعتمد التركيب الضوئي على انتشار ثنائي أكسيد الكربون من الهواء عبر المسام إلى الأنسجة المتوسطة. ويخرج الأكسجين، وهو منتج ثانوي من التركيب الضوئي، عبر المسام. يُفقد الماء بالتبخر حين تكون المسام مفتوحة ويجب تعويضه عبر تيار النتح بالماء الذي تمتصه الجذور. يجب أن توازن النباتات بين كمية ثنائي أكسيد الكربون الممتصة من الهواء وكمية الماء المفقودة عبر المسام، ويتحقق ذلك من خلال التحكم الفاعل والمنفعل بضغط امتلاء الخلايا الحارسة وحجم المسام.
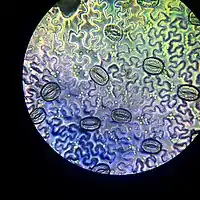
وظيفة الخلايا الحارسة
الخلايا الحارسة خلايا تحيط بالمسام. وتساعد على تنظيم معدل النتح عن طريق فتح وإغلاق المسام. الضوء هو المحفز الرئيسي لفتح المسام أو إغلاقها. للخلية الحارسة بشرة سميكة نسبيًا على الجانب المجاور للسُم ورقيقة في الجانب الآخر. حين يدخل الماء الخلية، ينتفخ الجانب الرقيق نحو الخارج بهيئة بالون ويسحب معه الجانب السميك، وتتخذ الخلية مظهر هلال؛ يشكل كل هلالين معًا فتحة سُم.[1]
تحوي الخلايا الحارسة بروتينات مستقبلة للضوء، وهي سيرين وكينازات ثريونين، تستقبل الضوء الأزرق. تشمل البروتينات المستقبلة للضوء نطاقين للضوء والأوكسجين والجهد، وهي جزء من عائلة نطاق بّي إيه إس الفائقة. تطلق البروتينات المستقبلة للضوء الكثير من الاستجابات مثل الانتحاء الضوئي وحركة الصانعات اليخضورية وتمدد الأوراق وفتح المسام. ولم يعرف الكثير عن آلية عمل هذه المستقبلات الضوئية قبل عام 1998. توضحت آلية عمل البروتينات المستقبلة للضوء من خلال التجارب على نبات الفول. أظهر الكشف المناعي واختبار اللطخة الغربية البعيدة أن الضوء الأزرق ينبه فوتوتروبين 1 وفوتوتروبين 2، ويبدأ عندها فوسفاتاز البروتين 1 شلال الفسفرة، الذي ينشط مضخة البروتون المعتمدة على الطاقة (إيه تي بّي)، وهي مضخة مسؤولة عن ضخ شوارد الهيدروجين خارج الخلية.[2] تسمح المضخة المفسفرة بارتباط بروتين 14-3-3 بنطاق مثبط ذاتي في المضخة عند نقطة النهاية سي. يُفسفر بعد ذلك السيرين والثريونين ضمن البروتين، وهو ما يفعّل نشاط مضخة شوارد الهيدروجين. وجدت التجربة ذاتها أنه عند الفسفرة، يرتبط بروتين 14-3-3 بالبروتينات المستقبلة للضوء قبل فسفرة المضخة. في تجربة مماثلة، توصل الباحثون إلى أن ارتباط البروتين 14-3-3 بموقع الفسفرة ضروري لتفعيل نشاط مضخة البروتون في الغشاء الخلوي. كشف ذلك من خلال إضافة الببتيدات الفوسفورية مثل بّي 950، التي تثبط ارتباط البروتين 14-3-3 بالمضخة المفسفرة وملاحظة تسلسل الحموض الأمينية. عندما تُضخ البروتونات نحو الخارج، تتجمع شحنة سلبية عبر الغشاء الخلوي. ويؤدي الاستقطاب المفرط للغشاء إلى تجمع شوارد البوتاسيوم وشوارد الكلور، فيزداد تركيز المُذاب وينخفض جهد الماء. يسمح الجهد السلبي للماء بحدوث التناضح في الخلية الحارسة، فيدخل الماء وتنتفخ الخلية.[3]
تتحكم التغيرات في ضغط الامتلاء في الخليتين الحارستين في فتح وإغلاق السُم. تتحكم حركة الشوارد والسكريات داخل وخارج الخلايا الحارسة بضغط الامتلاء فيها. جدار الخلايا الحارسة ذو سماكة متفاوتة (المنطقة الداخلية المجاورة للسُم أكثر سماكة وكوتينية جدًا) والألياف الدقيقة السيليلوزية مختلفة التوجه، ما يجعلها تنثني إلى الخارج عندما تكون منتفخة، فتسبب فتح المسام. تُغلق المسام عندما يُفقد الماء بالتناضح، الذي يحدث بسبب فقدان شوارد البوتاسيوم إلى الخلايا المجاورة.[4][5]
فقدان الماء وكفاءة استخدامه
الإجهاد المائي (الجفاف والإجهاد الملحي) واحد من المشاكل البيئية الرئيسية التي تسبب خسائر فادحة في الزراعة وفي الطبيعة. تساهم آليات متعددة في قدرة النباتات على تحمل الجفاف، منها ما يعمل من أجل استقرار النبات وحمايته من الأضرار الناجمة عن التجفف، والتحكم في كمية الخسارة المائية عبر المسام في أثناء الجفاف. يُنتج الهرمون النباتي المسمى حمض التسقيط (إيه بي أيه)، استجابة للجفاف.[6] وجرى التعرف على نوع رئيسي من مستقبلات إيه بي إيه. يسبب الهرمون النباتي إيه بي إيه إغلاق المسام استجابة للجفاف، ما يقلل من فقدان الماء عن طريق النتح ويسمح للنباتات بتجنب فقدان الماء في أثناء فترات الجفاف أو إبطائه. سيسمح استخدام نباتات زراعية قادرة على تحمل الجفاف بخفض خسائر الزراعة خلال فترات الجفاف. وبما أن الخلايا الحارسة تتحكم في فقدان النباتات للماء، فإن دراسة تنظيم فتح المسام وإغلاقها يمكن أن يساهم في تطوير نباتات أبطأ تأثرًا بالجفاف وأكثر كفاءة في استخدام الماء. وتبين دراسات أجراها جان بيير رونا أن إيه بي إيه هو مفتاح إغلاق المسام. إن تحفيزه يطلق شوارد البوتاسيوم والشوارد السالبة (الصواعد).[7][8] ويسبب تدفق الصواعد نزع استقطاب الغشاء الخلوي. يحفز نزع الاستقطاب شوارد البوتاسيوم الزائدة في الخلية لمغادرة الخلية بسبب عدم التوازن في جهد الغشاء. ويسبب هذا التغير المفاجئ في تركيز الشوارد تقلص الخلية الحارسة ويؤدي إلى إغلاق السم، وهو ما يقلل كمية الماء المفقود. تحدث هذه العمليات في سلسلة من التفاعلات وفقًا للبحث. وتسبب الزيادة في إيه بي إيه ارتفاع تركيز شوارد الكالسيوم. رغم الاعتقاد الأولي بأنها مجرد مصادفة، اكتشف لاحقًا أن الزيادة في شوارد الكالسيوم مهمة. ووجد أن شوارد الكالسيوم تشارك في تفعيل قنوات الصواعد. ما يسمح لها بالتدفق إلى الخلية الحارسة. وتشارك في منع مضخة البروتون من استعادة الاستقطاب. ولدعم فرضية دور الكالسيوم في كل هذه التغيرات الخلوية، صمم باحثون تجربة استخدموا فيها بروتينات تمنع إنتاج شوارد الكالسيوم. إذا كانت الفرضية صحيحة، ستقل التأثيرات التي يساهم فيها الكالسيوم عند استخدام البروتينات المثبطة. تبين أن الفرضية صحيحة إذ أدى استخدام المثبطات إلى ازدياد قدرة مضخة البروتون على استعادة الاستقطاب. ووُجد أيضًا أن تدفق الصواعد إلى داخل الخلايا الحارسة لم يكن بنفس القوة. هذان الأمران حاسمان في فتح المسام وإغلاقها ومنع فقدان الماء.[9]
مراجع
- Kinoshita, Toshinori؛ Emi, Takashi؛ Tominaga, Misumi؛ Sakamoto, Koji؛ Shigenaga, Ayako؛ Doi, Michio؛ Shimazaki, Ken-ichiro (01 ديسمبر 2003)، "Blue-Light- and Phosphorylation-Dependent Binding of a 14-3-3 Protein to Phototropins in Stomatal Guard Cells of Broad Bean"، Plant Physiology، 133 (4): 1453–1463، doi:10.1104/pp.103.029629، ISSN 0032-0889، PMC 300702، PMID 14605223.
- Imamura S (1943) Untersuchungen uber den mechanismus der turgorschwankung der spaltoffnungs-schliesszellen. Jap. J. Bot. 12:251-346.
- Kinoshita, Toshinori؛ Shimazaki, Ken-ichiro (15 نوفمبر 2002)، "Biochemical Evidence for the Requirement of 14-3-3 Protein Binding in Activation of the Guard-cell Plasma Membrane H+-ATPase by Blue Light"، Plant and Cell Physiology، 43 (11): 1359–1365، doi:10.1093/pcp/pcf167، ISSN 1471-9053، PMID 12461136.
- Humble GD & Raschke K (1971) Stomatal opening quantitatively related to potassium transport. Evidence from electron probe analysis. Plant Physiol. 48:447-453.
- Schroeder JI, Hedrich R, & Fernandez JM (1984) Potassium-selective single channels in guard cell protoplasts of Vicia faba. Nature 312:361-362.
- Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow TF, Alfred SE, Bonetta D, Finkelstein R, Provart NJ, Desveaux D, Rodriguez PL, McCourt P, Zhu JK, Schroeder JI, Volkman BF, & Cutler SR (2009) Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324:1068-1071.
- Schroeder JI, Kwak JM, & Allen GJ (2001) Guard cell abscisic acid signaling and engineering drought hardiness in plants. Nature 410:327-330.
- Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, & Grill E (2009) Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 324:1064-1068.
- Meimoun, Patrice؛ Vidal, Guillaume؛ Bohrer, Anne-Sophie؛ Lehner, Arnaud؛ Tran, Daniel؛ Briand, Joël؛ Bouteau, François؛ Rona, Jean-Pierre (سبتمبر 2009)، "Intracellular ca2+ stores could participate to abscisic acid-induced depolarization and stomatal closure in Arabidopsis thaliana"، Plant Signaling & Behavior، 4 (9): 830–835، doi:10.4161/psb.4.9.9396، ISSN 1559-2316، PMC 2802785، PMID 19847112.
 بوابة علم النبات
بوابة علم النبات