صفيحة قاعية
الصفيحة القاعية (floor plate) هي بنية تشكل جزءًا من الجهاز العصبي النامي للكائنات الفقارية. بوقوعها على الخط الناصف البطني لأنبوب العصبي الجنيني، تُعد الصفيحة القاعيية بنية دبقية متخصصة تعبر المحور الأمامي الخلفي من المخ الأوسط إلى مناطق المؤخرة. وقد تبين أن الصفيحة القاعية لا تزال موجودة بين الفقاريات مع وجود بنى مماثلة في اللافقاريات مثل ذبابة الفاكهة الدورسيلا والديدان الأسطوانية الخيطية. ومن الناحية الوظيفية، تعد هذه البنية بمثابة المنظم للأنسجة البطنية في الجنين إضافة إلى وظيفتها في توجيه عملية تحديد مواقع الخلايا العصبية والتمايز على طول المحور الظهراني البطناني للأنبوب العصبي.[1][2][3]
| صفيحة قاعية | |
|---|---|
| تفاصيل | |
| جزء من | أنبوبة عصبية |
| UBERON ID | 0003079 |
التأثير
تم دراسة تأثير الصفيحة القاعية أثناء مرحلة التطور الجنيني لأجنة الفقاريات باستفاضة في الكتكوت وسمك الزرد ويحدث هذا التأثير نتيجة لوجود شبكة إشارات معقدة بين الأنسجة، ولكن لم يتم تنقيح تلك التفاصيل بالكامل حتى الآن. وهناك حاليًا العديد من مدارس الفكر المتنافسة فيما يخص هذا الأمر. أولاً قد يتم توسيط تمايز الصفيحة القاعية عن طريق الإشارة الاستقرائية من الحبل الظهري المستبطن، وهو عبارة عن نسيج إشارة مشتق من الأديم المتوسط المحوري. وتم تدعيم ذلك من الناحية التجريبية في الكتكوت، الذي تم فيه حث تأثير الصفيحة القاعية، إضافة إلى تمايز النسيج العصبي البطني الترابطي، وتوسيطه عن طريق جزيئات الإشارة التي تم إفرازها القنفذ سونيك. ويتم التعبير عن هذه الجزيئات في مقياس مدرج مائل مع وضع أعلى تركيز في الحبل الظهري والصفيحة القاعية. تُظهر تجارب التطعيم في المختبر أن إزالة هذا الجزيء يمنع تمايز الصفيحة القاعية، في حين أن إفرازها المنتبذ يؤدي إلى تمايز خلاياها.[4] وترجح وجهة نظر أخرى أن جذع خلايا الصحيفة القاعية للأنبوبة العصبية الناتجة من الخلايا الناشئة من الأديم المتوسط المحوري. وفيما يتعلق بالتجارب المختلطة بين الكتكوت والسمان إضافة إلى تجارب التفاعل الوراثي في سمك الزرد، بدا أن خلايا الحبل الظهري والصحيفة القاعية تنشأ من طلائع مشتركة. وهنا تكون الإشارة العقدية مطلوبة لتمايز خلايا الصفيحة القاعية المتوسطة في سمك الزرد، في حين أن القنفذ سونيك قابل للتمدد. تشير هذه البيانات إلى أن آلية حث الصفيحة القاعية تختلف في الحيوانات السلوية عن الحيوانات اللاسلوية.[5] ولتوفيق هذه الفروق، تم اقتراح نموذج حث مزدوج في الكتكوت. وتم حث الخلايا المتعلقة بالأديم الظاهر بكثافة في هذا النموذج لتصبح صفيحة قاعية أثناء مرحلة المعيدة عن طريق الأديم المتوسط القردودي، وربما عن طريق الإشارة العقدية. وفيما بعد في النمو أثناء تكون العصيبة، يؤدي التواصل والتفاعل الموسع بين خلايا الحبل الظهري والصفيحة القاعية إلى حدوث تمايز، مما يدل على التأثير المشترك بين الإشارة العقدية وإشارة القنفذ سونيك.[6]
توجيه المحور العصبي
يُعد قرار عبور العصبونات للخط الناصف في تطور الجهاز العصبي المركزي أو عدم العبور أمرًا بالغ الأهمية. ويتم توسيط هذا الاختيار في الفقاريات عن طريق الصفيحة القاعية، ويتمكن الجنين من تطوير النسيج العصبي في نصفي الجسم الأيمن والأيسر بنجاح، على سبيل المثال، ولكن العصبونات الموجودة على نفس الجانب لا تعبر الخط الناصف، حيث إن العصبونات الصوارية تعبر الخط الناصف وتشكل صوارًا واحدًا. تنمو هذه العصبونات الخاصة في المنطقة الظهرية من الأنبوب العصبي وتنتقل بطنيًّا نحو الصفيحة القاعية. وعند الوصول إلى الصفيحة القاعية، تعبر الخلايا العصبية الصوارية من خلال الأنسجة لتظهر في الجانب المقابل للأنبوب العصبي، ومن ثم تنتأ إما في الناحية الأمامية أو الخلفية في الأنبوب.[7]
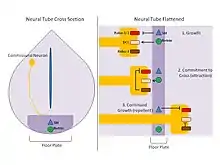
- النترينات: النترينات عبارة عن بروتينات تخرج وتُفرز عن طريق خلايا الصفيحة القاعية. ثبت من خلال التجارب التي تستخدم مستخلصات الصفيحة القاعية والخلايا العصبية الصوارية الثابتة في مصفوفة الكولاجين انجذاب الخلايا العصبية نحو الصفيحة القاعية في المختبر.[8] إضافة إلى ذلك، فإن فصل وتعداء نترين 1 ونترين 2، وهما اثنان من البروينات المفرزة في خلايا الرأس يسفر عن تأثيرات مماثلة .[9] أكدت بحوث أخرى أن النترينات تعمل كبروتينات جاذبة مع القنفذ سونيك لتوجيه المحاور العصبية الصوارية نحو الصفيحة القاعية.[10] تُفرز النترينات من قبل خلايا الصفيحة القاعية ووظيفتها ربط مستقبلات DCC في حالة الانجذاب الكيميائي.
- فَلْعَة: فَلْعَة عبارة عن لجين يتم إفرازه في الصفيحة القاعية ويعمل على منع عبور المحور العصبي من خلال الأنبوب العصبي. بينما تجذب النترينات الخلايا العصبية الصوارية تجاه الخط الناصف، تصد بروتينات الفلعة الخلايا العصبية وتطردها من الخط الناصف. وبما أن المحاور العصبية غير قادرة على عبور خط المنتصف عن طريق الأنبوبة العصبية، فستُصد عن طريق لجينات الفلعة المستخرجة من خلايا الصفيحة القاعية. تعمل الفلعة عن طريق مستقبلاتها المستديرة (روبو) 1 و2. ويعيق هذا التفاعل الانجذاب الكيميائي المقدم عن طريق النترينات/دي سي سي. ومع ذلك، يتم تنظيم روبو 3 (1) تصاعديًا أثناء نمو المحاور العصبية الصوارية أثناء الانتقال تجاه الصفيحة القاعية، التي تحتجز روبو 1 و2 في الخلية داخل الحويصلة. ومن ثم، فإن مسار جذب النترينات/دي سي سي يسيطر على مسار إبعاد الفلعة/روبو وقد ينمو المحور العصبي تجاه الخط الناصف ويدخل الصفيحة القاعية. وعند الدخول، عن طريق آلية لا تزال غير مفهومة تمامًا حتى الآن، يصبح تنظيم روبو 3 تنازليًّا وهذا يحرر روبو 1 و2 ويسمح بالتنظيم التصاعدي لهما، ويتصدى للعصبون بفاعلية من الخط الناصف للصفيحة القاعية. وعن طريق هذا العبور المعقد للفلعة، يتم توجيه روبو 1 وروبو 2 والمحاور العصبية الصوارية تجاه الخط الناصف لعبور الأنبوبة العصبية ومنعها من العبور ثانية.[11]
وقد درست جزيئات الإشارة التي توجه نمو المحاور العصبية الصوارية وبروزها في اللافقاريات جيدًا. في مسار الجذب الكيميائي للنترين ودي سي سي، تعد متشابهات الربداء الرشيقة هي يو إن سي 6/يو إن سي 40 ويو إن سي 5 بينما تعدد متشابهات ذبابة الفاكهة نترين أ ونترين ب/ أنهك ودونك 5. وفي مسار إبعاد الفلعة/روبو تعد متشابهات الربداء الرشيقة إس إل تي 1 وساكس 3 بينما تعد متشابهات ذبابة الفاكهة معروفة أيضًا على أنها الفلعة/روبو (1-3).[7]
مراجع
- "Wolpert, Lewis. Principles of Development: 3rd Edition. Oxford University Press, 2007."
- "Gilbert, Scott F. Principles of Development: 8th Edition. Sinauer Associates, Inc. 2006."
- "Jessell, Thomas M. Neuronal Specification in the spinal cord: inductive signals and transcriptional codes. Nature Reviews Genetics. Oct, 2000(1)"
- "Yamada, T. Control of Cell Pattern in the Developing Nervous System: Polarizing Activity of the Floor Plate and Notochord. Cell, Vol. 64, 635-647, February 8, 1991"
- "Strahle, Uwe, et al. Vertebrate floor-plate specification: variations on common themes. Trends in Genetics Vol.20 No.3 March 2004"
- "Patten, Iain, et al. Distinct modes of floor plate induction in the chick embryo. 19 June 2003. Development 130, 4809-4821"
- "Guan KL and Rao Y. Signalling mechanisms mediating neuronal responses to guidance cues. Nat Rev Neurosci. 2003 Dec;4(12):941-56."
- "Serafini, Tito, et al. The Netrins Define a Family of Axon Outgrowth-Promoting Proteins Homologous to C. elegans UNC-6. Cell, Vol. 79, 409-424. August 12, 1994"
- "Kennedy, Timothy E, et al. Netrins Are Diffusible Chemotropic Factors for Commissural Axons in the Embryonic Spinal Cord. Cell, Vol. 79, 425-435, August 12, 1994"
- "Charron, F, et al. The morphogen sonic hedgehog is an axonal chemoattractant that collaborates with netrin-1 in midline axon guidance. Cell. 2003 Apr 4;113(1):11-23."
- "Long, Hua et al. Conserved Roles for Slit and Robo Proteins in Midline Commissural Axon Guidance. Neuron, Vol. 42, 213–223, April 22, 2004"
 بوابة طب
بوابة طب بوابة علم الأحياء
بوابة علم الأحياء