2-succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate synthase
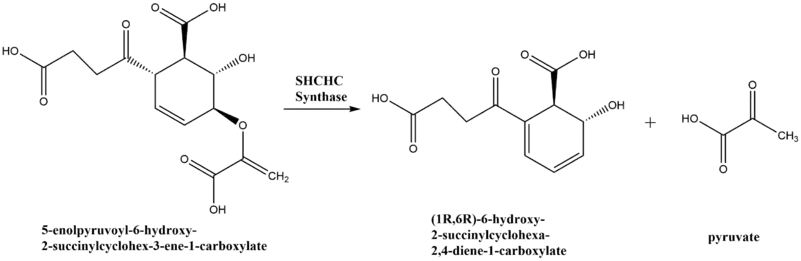
2-Succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate synthase (EC 4.2.99.20), also known as SHCHC synthase is encoded by the menH gene in Escherichia coli and functions in the synthesis of vitamin K.[1] The specific step in the synthetic pathway that SHCHC synthase catalyzes is the conversion of 5-enolpyruvoyl-6-hydroxy-2-succinylcyclohex-3-ene-1-carboxylate to (1R,6R)-6-hydroxy-2-succinylcyclohexa-2,4-diene-1-carboxylate and pyruvate.[2]
| 2-succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate synthase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC no. | 4.2.99.20 | ||||||||
| CAS no. | 122007-88-9 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Background
Vitamin K is a fat soluble vitamin known to aid in blood clotting. It is recommended that all newborns receive an injection of vitamin K in order to prevent excessive bleeding of the brain after birth. There are two major forms of vitamin K that occur naturally. Phylloquinone, also known as K1, is synthesized by plants and is the major form of vitamin K in the diet. Menaquinone, K2, includes a range of forms that are synthesized by bacteria in the gut.[3]
Vitamin K is synthesized from the molecule chorismate in a nine step conversion process. SHCHC synthase catalyzes the third step in the process.[4]
Chemistry
Reaction scheme
Enzyme Structure

The crystal structure of the MenH enzyme in E.coli (SHCHC synthase) exists as a complex of three protein molecules shown in the diagram. SHCHC synthase forms an alpha/beta hydrolase fold with a central set of seven parallel beta sheets surrounded by alpha helixes on both sides. A cap of five alpha helixes serves to enclose the active site.[5] The enzyme exists in an open form until it binds the substrate, when it morphs into a closed form with an active catalytic triad.[6]
Energetic analysis shows that SHCHC synthase has a low energetic burden for catalytic activity.[1] This means the enzyme is more prone to mutation and is one of the most diverse enzymes in the vitamin K synthetic pathway.[7] Only fifteen amino acid residues are absolutely conserved across mutations of the enzyme.[7]
Catalytic Mechanism
The active site contains a catalytic triad of syrine, histine and arginine, which is conserved across all mutants and is proposed to initiate the reaction.[1] The triad residues are located at Ser86, Asp210, and His232.[5] This triad is proposed to catalyze a proton extraction which triggers a transfer of electrons leading to the elimination of pyruvate and formation of SHCHC.[6] Originally, it was proposed that the transition state was stabilized by a nontraditional oxyanion hole. Now a traditional oxyanion hole is favored, but not definitive.[5]
Reaction Mechanism

Controversy
MenH (SHCHC synthase) was previously thought to be a thioesterase involved in hydrolyzing DHNA-CoA in a later step of menaquinone synthesis. In 2008, it was determined that MenH has poor catalytic activity toward palmitoyl-CoA, casting doubt on its role as a thioesterase.[1] Direct analysis confirmed that MenH is unable to hydrolyze DHNA-CoA.[1] In 2009, it was proposed that a dedicated hotdog fold thioesterase would be needed to catalyze the hydrolysis of DHNA-CoA.[8] A protein was identified in 2013 that could fit this role.[9]
References
- Jiang M, Chen X, Guo ZF, Cao Y, Chen M, Guo Z (March 2008). "Identification and characterization of (1R,6R)-2-succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate synthase in the menaquinone biosynthesis of Escherichia coli". Biochemistry. 47 (11): 3426–34. doi:10.1021/bi7023755. PMID 18284213.
- "Information on EC 4.2.99.20 - 2-succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate synthase". Brenda: The Comprehensive Enzyme Information System. TU Braunschweig. 2014-07-01. Retrieved 2014-11-01.
- "Micronutrient Information Center". Linus Pauling Institute. 2014-11-30. Retrieved 2014-11-30.
- van Oostende C, Widhalm JR, Furt F, Ducluzeau AL, Basset GJC (2011) Phylloquinone (Vitamin K1): function, enzymes and genes. in Advances in Botanical Research, eds Fabrice Rébeillé and Roland Douce, 59: 229-61, Academic Press (Amsterdam).
- PDB: 4GDM; Johnston JM, Jiang M, Guo Z, Baker EN (2013-04-18). "Crystal structures of E. coli native MenH and two active site mutants". PLOS ONE. 8 (4): e61325. Bibcode:2013PLoSO...861325J. doi:10.1371/journal.pone.0061325. PMC 3630204. PMID 23637813.
- Sun Y, Yin S, Feng Y, Li J, Zhou J, Liu C, Zhu G, Guo Z (May 2014). "Molecular basis of the general base catalysis of an α/β-hydrolase catalytic triad". The Journal of Biological Chemistry. 289 (22): 15867–79. doi:10.1074/jbc.M113.535641. PMC 4140940. PMID 24737327.
- Jiang M, Chen X, Wu XH, Chen M, Wu YD, Guo Z (July 2009). "Catalytic mechanism of SHCHC synthase in the menaquinone biosynthesis of Escherichia coli: identification and mutational analysis of the active site residues". Biochemistry. 48 (29): 6921–31. doi:10.1021/bi900897h. PMID 19545176.
- Widhalm JR, van Oostende C, Furt F, Basset GJ (April 2009). "A dedicated thioesterase of the Hotdog-fold family is required for the biosynthesis of the naphthoquinone ring of vitamin K1". Proceedings of the National Academy of Sciences of the United States of America. 106 (14): 5599–603. Bibcode:2009PNAS..106.5599W. doi:10.1073/pnas.0900738106. PMC 2660889. PMID 19321747.
- Chen M, Ma X, Chen X, Jiang M, Song H, Guo Z (June 2013). "Identification of a hotdog fold thioesterase involved in the biosynthesis of menaquinone in Escherichia coli". Journal of Bacteriology. 195 (12): 2768–75. doi:10.1128/JB.00141-13. PMC 3697248. PMID 23564174.