Alteromonas macleodii
Alteromonas macleodii is a species of widespread marine bacterium found in surface waters across temperate and tropical regions.[1] First discovered in a survey of aerobic bacteria in 1972, A. macleodii has since been placed within the phylum Pseudomonadota and is recognised as a prominent component of surface waters between 0 and 50 metres.[2][3][4] Alteromonas macleodii has a single circular DNA chromosome of 4.6 million base pairs.[5] Variable regions in the genome of A. macleodii confer functional diversity to closely related strains and facilitate different lifestyles and strategies.[1][6] Certain A. macleodii strains are currently being explored for their industrial uses, including in cosmetics,[7] bioethanol production and rare earth mining.[9]
| Alteromonas macleodii | |
|---|---|
| Scientific classification | |
| Domain: | |
| Phylum: | |
| Class: | |
| Order: | |
| Family: | |
| Genus: | |
| Binomial name | |
| Alteromonas macleodii Baumann et al., 1972 | |
| Synonyms | |
|
Alteromonas macleodii | |
Morphology
The species Alteromonas macleodii refers to an encapsulated gram-negative heterotrophic γ-proteobacterium.[10][11] It is aerobic and motile, with a singular unsheathed polar flagellum.[1] Isolates of A. macleodii are between 0.6 to 0.8 μm width and 1.4 to 2.0 μm length, and are neither luminescent nor pigmented. Alteromonas macleodii is able to grow on glucose-only solid medium, forming colonies up to 0.9 cm in diameter with irregular edges.[11] As a result of phenotypic variability and differences in genomic content among strains, competitiveness in culture varies both between cultures of the same strain and between strains from different geographical areas and depths.[6]
Bacteria classified as A. macleodii are r-strategists; large cells with high nucleic-acid content are commonly seen, with high dividing frequencies and carbon production rates.[12] As a copiotroph, A. macleodii is able to use glucose as its sole carbon and energy source and blooms under high nutrient and sodium concentrations where it is able to outcompete other organisms. Low temperatures and low carbon availability generally impede growth.[1]
Distribution

Alteromonas macleodii are ubiquitous in the global oceans, typically adhering to small organic particles in the upper 50 metres of the water column.[13] They constitute a significant proportion of the bacterial abundance in the North Atlantic and Mediterranean at up to 9 and 23 percent of total particle-attached bacteria respectively,[10] and are also present in the Northeast Pacific and subtropical Atlantic.[14]
Initially, two ecotypes of Alteromonas macleodii were described, as niche differentiation had caused two distinct strains of the bacterium to occupy different water depth profiles. The “deep ecotype” is more suited to microaerophilic environments and it sinks rapidly into the deeper pelagic zones, relying on a different spectrum of carbon sources.[13][15] Recently, the deep ecotype strains have been reclassified as Alteromonas mediterranea.[16]
Physiology
Physiological variation in Alteromonas macleodii leads to specific adaptive strategies in terms of carbon and iron metabolism, cellular communication, and nutrient acquisition.[1] Some strains are specialised in their associations with prochlorococcus species through enhanced phenol degradation, while others have a unique capacity to metabolise sugars from specific algae species. The production of homoserine, lactones and siderophores are also strain-specific, with different nutrient acquisition and cellular communication strategies between strains under different ecological conditions.[6]
The physiology of Alteromonas macleodii can influence iron concentrations and recalcitrant dissolved organic matter (DOM) production in the oceans. These bacteria utilise unique Ton-B dependent transporters to acquire iron as well as carbon substrates. As a result, some strains of A. macleodii are able to more efficiently regulate the uptake of glucose, tryptophan, and tyrosine during growth.[17] An ATPase-independent mechanism is involved in the transport of the siderophores which scavenge iron for the bacterium, iron which is then used by enzymes to facilitate carbon metabolism.[17][18]
The physiological responses of A. macleodii depend on the type of amino acids taken up. D-amino acids such as D-alanine, D-serine and D-glutamic acid reduce metabolic activity, also inhibiting the production of exopolysaccharide (EPS).[19] Exopolysaccharide production contributes to cell aggregation and the formation of biofilms in marine bacterial species.[20] Paradoxically, the uptake of D-amino acids by A. macleodii impedes the production of EPS, but encourages the formation of biofilms by promoting other independent aggregation factors.[19]
The extracellular membrane vesicles in A. macleodii play a crucial role in algae degradation and habitat colonisation. These vesicles contain hydrolytic enzymes such as lipases, proteases and nucleases. These enzymes are responsible for the degradation of cell walls and inner components of red algae such as Kappaphycus. Alteromonas macleodii is also very efficient at degrading alginate, expressing as many as five separate alginate lyases and outcompeting other bacterial groups when grown on this substrate.[21][22]
Ecology
The surface-dwelling A. macleodii is well-suited to the degradation of a variety sugars and amino acids. These bacteria are generally attached to small particles, but can also be free-living and are able to utilise a number of different substrates for growth.[6][21][23] Alteromonas macleodii is a copiotroph flexible in its use of substrates, growing rapidly at high carbon and nutrient concentrations.[1]
Interspecies interactions
Alteromonas macleodii is able to sustain Prochlorococcus cells undergoing chlorosis due to nutrient stress. During chlorosis, Prochlorococcus is unable to use its essential photosynthetic pigments, but is able to survive for an extended period of time in the presence of A. macleodii.[24] The marine heterotroph has also been found associated with Trichodesmium, a filamentous cyanobacteria that fixes nitrogen in the oceans. Alteromonas macleodii might influence Trichodesmium metabolism, allowing for the catabolism of methanol and the detoxification of radical oxygen species.[25]
Iron limitation
Under iron replete conditions, the rate of respiration in A. macleodii is significantly reduced. Iron metal is associated with several key processes for bacterial metabolism, such as the citric acid cycle, glycolysis, and oxidative phosphorylation, all of which are functionally limited when iron availability is not sufficient. The growth rate of A. macleodii is therefore reduced when iron is limited, although the growth rate of strains from coastal populations is reduced more so than those from mid-oceanic populations.[26]
Copper stress
Increased exposure of bacteria to copper may occur in several ways, such as nutrient leaching, metals from ship hulls, or natural mineral deposits. Under conditions of increased copper concentrations, biofilm production of A. macleodii significantly increases as a defensive response to copper induced stress. These bacteria are able to colonise areas of very high copper concentration, giving them an advantage over other bacteria under these conditions.[27][28]
Role in carbon cycle
As heterotrophic bacteria, A. macleodii consume dissolved organic carbon in seawater and are then consumed by higher trophic levels, acting as a gateway for carbon into ecosystems. While natural ecosystems consist of a variety of heterotrophs contributing to the carbon cycle, it has been found in laboratory settings that A. macleodii is capable of drawing down the complete pool of labile DOC present in coastal waters. This indicates that the relationship between A. macleodii and other bacteria in the microbial loop of coastal waters is one of functional redundancy: Alteromonas is capable of carbon cycling to the same extent as entire microbial communities.[12]
Cell wall polysaccharides secreted by macroalga are degraded by microbes such as Alteromonas, and are a major source of carbon into marine ecosystems. Alteromonas macleodii exhibits two distinct strategies for carbon uptake, depending on the type of polysaccharide present in their habitat. When degrading the common polysaccharides laminarin, alginate, and pectin, A. macleodii releases different catabolites at different times to degrade the respective substrates. Laminarin is the first polysaccharide that is degraded, followed by alginate and pectin. This temporal variation in carbon utilisation is a result of a shift in transcriptional activity of CAZymes and polysaccharide utilisation gene fragments. The biphasic nature of these cellular adaptations indicates that A. macleodii’s role in the drawdown of polysaccharide DOC is adaptable to changing community structures of macroalgal communities.[29]
Alginate is a gel textured polysaccharide that is a common component of macroalgal cell walls, and is a nutrient and carbon source for many organisms. A. macleodii are a key components of the carbon cycle, in that they degrade alginate, increasing DOC drawdown in marine environments. Further, A. macleodii has been found to outcompete other species of bacteria in the degradation of alginate, indicating that A. macleodii plays a particularly relevant role in ecological carbon cycling.[30] The degradation of algal polysaccharides and proteins is crucial for nutrient acquisition, and has the effect of preventing overgrowth of red algae.
Genome
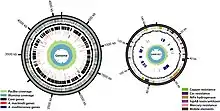
The genomes of A. macleodii strains have distinct genomic content associated with different lifestyles and geographical locations.[23][6] Small differences in overall nucleotide identity between strains can be functionally substantial; many important functional genes are found on genomic islands (GIs) exchanged between populations.[31][32] Strains associated with surface waters such as ATCC 21726 have a single circular genome of about 4.6 million base pairs.[5] Alteromonas macleodii has an estimated 4400 total genes[6] with about 47% GC content.[5] Its pangenome is vast,[13] there is significant variation in functional genetic content between strains from different geographical regions.[4] Additionally, closely related strains vary in functional genes found on genomic islands.[1][6][33]
Alteromonas macleodii is globally distributed in the surface ocean at 0-50m depth,[1][13] these strains are highly variable functionally despite sharing 97-99% nucleotide identity. Functional differences between surface strains are conferred by horizontally transferred genes, and are reflective of the variable conditions of surface waters.[1][6] Surface strains of A. macleodii also have a higher number of genes associated with utilising different sugar and amino acid substrates as well as transcriptional regulators for plasticity in changing conditions.[23] This plasticity is associated with the ability of Alteromonas to grow rapidly and take advantage of increases in available organic matter.[34] Key genes associated with siderophore production and degradation of algal substrates are also transferred horizontally.[6][35]
Genomic Islands
Content of genomic islands differs greatly between strains, especially those coding for polysaccharides that present on the flagellum and the outer surface of the cell, with possible roles in phage avoidance.[1] Some strains have acquired heavy-metal tolerance and other important functional genes from GIs found in A. mediterranea,[31] GIs containing important functional genes are exchanged between different populations of A. macleodii, increasing functional flexibility.[32] Plasmids carried by some A. macleodii strains that enhance heavy-metal tolerance are found in genomic islands in other members of the Alteromonadales.[6]
Heavy metal tolerance
Alteromonas macleodii is an early coloniser of copper-based antifouling paint on ships, where it forms biofilms.[36][28] While there is variability between strains, A. macleodii generally has genomic features which confer very high tolerance to copper and other heavy metals.[6] Strains with high copper tolerance all had at least one genomic island with metal tolerance genes, including several copies of the key cytoplasmic detoxifying factor copA and its transcriptional regulator merR. These factors give A. macleodii the ability to grow at metal levels lethal to most other marine species.[28][37] Megaplasmids found in particularly metal-tolerant strains contain multiple copies of metal detoxification systems with orthologs in Escherichia and Pseudomonas.[38]
Copper prevents bacterial growth due to its intrinsic antimicrobial properties, minimising the formation of biofilms.[39] However, some A. macleodii strains are still able to induce growth of biofilms under elevated copper concentrations. These strains possess an alteration in the diguanylate cyclase (DGC) genes, which control the expression of biofilms in A. macleodii.[40] Specific subsets of the DGC genes are highly expressed in some strains, enhancing biofilm development by amplifying the transduction of signals that promote biofilm formation. This process changes the structure of the microbial community, affecting both the microenvironment and biogeochemical cycling.[40]
Relationship to Alteromonas medditerranea
A closely related set of strains previously considered "deep-ecotype" of A. macleodii have since been reclassified under A. mediterranea as they share only 81% overall sequence identity.[16] There are 3200 genes shared between A. macleodii and A. mediterranea, with 1200-1600 unique to each.[23] The "deep-ecotype" A. mediterranea strains contain more dioxygenases for degrading recalcitrant DOM such as urea, molecular chaperones for protein folding at lower temperatures and hydrogenases associated with heavy-metal tolerance, located with other tolerance genes on a single GI.[23] These sets of genes are not exclusive to A. mediterranea, as they are exchanged between different populations of Alteromonas species along with other sets of functional genes such as enzymes for sugar and amino acid degradation, allowing for niche specialisation.[10][33]
Phage infecting A. macleodii
Genes such as phase integrases and the CRISPR cluster found in some A. macleodii strains are likely involved in phage interactions.[23] Some genomic islands encode specific surface receptors recognised by phages, increasing the susceptibility to phage infections.[41] Some components of GIs are lysogenic or defective phages; one of these widespread GIs encodes virus-derived mismatch repair and RNA chaperone genes.[1] For example, the O-chain lipopolysaccharides of the bacteria may associate with the receptor binding proteins present on the Alterophage R8W, allowing attachment and entry.[41] Just over a dozen Alterophages infecting A. macleodii have been characterised to date, including members of the Podoviridae[42] and Autographiviridae.[43]
Industrial Uses
Reduction of potassium tellurite to elemental tellurium
Elemental tellurium is an extremely rare metalloid contained in the Earth's crust with desirable optic and electronic properties.[44] However, current industrial production of tellurium requires the usage of substances harmful to both humans and the environment. As a result, extraction of metalloids by biotechnological applications involving bacterial biosynthesis of nanoparticles from various uncommon and rare metals are increasingly being studied.[9]
Tellurium has toxic effects on bacteria through an unknown mechanism. Alteromonas macleodii contains a plasmid that houses genes allowing for resistance to multiple metals, and has the ability to reduce potassium tellurite into elemental tellurium. The nanoparticles of the reduced tellurium are diffused into the cytoplasm, or into the extracellular space in the form of both electron-dense globules and metalloid crystals.[9] This makes A. macleodii a candidate for facilitating the extraction of tellurium with reduced reliance on toxic chemicals.
Extraction of biomolecules from red seaweeds
Membrane vesicles containing κ-carrageenase are produced by A. macleodii, which allows it to degrade carrageenan, a major polysaccharide found in the cell walls of red seaweeds.[45] The κ-carrageenase containing vesicles can be exploited for bioethanol production since they convert carbohydrate-rich biomass to sugars. Biomolecules present in the red seaweeds, such as vitamins and carotenoids, are also extracted for commercial use in tandem with the bioethanol production process.
EPS deepsane usage in cosmetics
Alteromonas macleodii secretes "deepsane", an exopolysaccharide now used in cosmetics.[7][46] Studied properties of "deepsane" include high viscosity possibly due to the interaction between acetate and pyruvate, making it an alternative to other viscous polymers currently used in food and cosmetics.[7] As of 2012, "deepsane" is also commercially available in the cosmetics industry and is referred to as Abyssine®, used in skincare products to reduce skin irritation from sunburns.[47]
Additional future prospects in water treatments
Alteromonas macleodii, when grown in glucose-supplemented media, secreted an unexpectedly high-molecular-weight polymer that changed carbohydrate composition. The polymer was found to be rich in uronic acids and therefore expected to have a heavy-metal-binding ability that could be used and applied in the treatment of wastewater.[11]
References
- López-Pérez M, Gonzaga A, Martin-Cuadrado AB, Onyshchenko O, Ghavidel A, Ghai R, Rodriguez-Valera F (2012). "Genomes of surface isolates of Alteromonas macleodii: the life of a widespread marine opportunistic copiotroph". Scientific Reports. 2 (1): 696. Bibcode:2012NatSR...2E.696L. doi:10.1038/srep00696. PMC 3458243. PMID 23019517.
- Baumann L, Baumann P, Mandel M, Allen RD (April 1972). "Taxonomy of aerobic marine eubacteria". Journal of Bacteriology. 110 (1): 402–429. doi:10.1128/jb.110.1.402-429.1972. PMC 247423. PMID 4552999.
- Floyd MM, Tang J, Kane M, Emerson D (June 2005). "Captured diversity in a culture collection: case study of the geographic and habitat distributions of environmental isolates held at the american type culture collection". Applied and Environmental Microbiology. 71 (6): 2813–2823. Bibcode:2005ApEnM..71.2813F. doi:10.1128/aem.71.6.2813-2823.2005. PMC 1151842. PMID 15932972.
- Garcia-Martinez, Jesus; Acinas, Silvia G.; Massana, Ramon; Rodriguez-Valera, Francisco (2002). "Prevalence and microdiversity of Alteromonas macleodii-like microorganisms in different oceanic regions". Environmental Microbiology. 4 (1): 42–50. doi:10.1046/j.1462-2920.2002.00255.x. ISSN 1462-2912. PMID 11966824.
- Gonzaga, Aitor; López-Pérez, Mario; Martin-Cuadrado, Ana-Belen; Ghai, Rohit; Rodriguez-Valera, Francisco (2012). "Complete Genome Sequence of the Copiotrophic Marine Bacterium Alteromonas macleodii Strain ATCC 27126". Journal of Bacteriology. 194 (24): 6998. doi:10.1128/jb.01565-12. ISSN 0021-9193. PMC 3510622. PMID 23209244.
- Koch H, Germscheid N, Freese HM, Noriega-Ortega B, Lücking D, Berger M, et al. (January 2020). "Genomic, metabolic and phenotypic variability shapes ecological differentiation and intraspecies interactions of Alteromonas macleodii". Scientific Reports. 10 (1): 809. Bibcode:2020NatSR..10..809K. doi:10.1038/s41598-020-57526-5. PMC 6972757. PMID 31964928.
- Raguénès G, Cambon-Bonavita MA, Lohier JF, Boisset C, Guezennec J (June 2003). "A novel, highly viscous polysaccharide excreted by an alteromonas isolated from a deep-sea hydrothermal vent shrimp". Current Microbiology. 46 (6): 448–452. doi:10.1007/s00284-002-3922-3. PMID 12732953. S2CID 23929160.
- Beleneva IA, Efimova KV, Eliseikina MG, Svetashev VI, Orlova TY (September 2019). "The tellurite-reducing bacterium Alteromonas macleodii from a culture of the toxic dinoflagellate Prorocentrum foraminosum". Heliyon. 5 (9): e02435. doi:10.1016/j.heliyon.2019.e02435. PMC 6819836. PMID 31687549.
- García-Martínez J, Acinas SG, Massana R, Rodríguez-Valera F (January 2002). "Prevalence and microdiversity of Alteromonas macleodii-like microorganisms in different oceanic regions". Environmental Microbiology. 4 (1): 42–50. doi:10.1046/j.1462-2920.2002.00255.x. PMID 11966824.
- Raguenes G, Pignet P, Gauthier G, Peres A, Christen R, Rougeaux H, et al. (January 1996). "Description of a new polymer-secreting bacterium from a deep-sea hydrothermal vent, Alteromonas macleodii subsp. fijiensis, and preliminary characterization of the polymer". Applied and Environmental Microbiology. 62 (1): 67–73. Bibcode:1996ApEnM..62...67R. doi:10.1128/aem.62.1.67-73.1996. PMC 167774. PMID 8572714.
- Pedler BE, Aluwihare LI, Azam F (May 2014). "Single bacterial strain capable of significant contribution to carbon cycling in the surface ocean". Proceedings of the National Academy of Sciences of the United States of America. 111 (20): 7202–7207. Bibcode:2014PNAS..111.7202P. doi:10.1073/pnas.1401887111. PMC 4034236. PMID 24733921.
- Ivars-Martínez E, D'Auria G, Rodríguez-Valera F, Sânchez-Porro C, Ventosa A, Joint I, Mühling M (September 2008). "Biogeography of the ubiquitous marine bacterium Alteromonas macleodii determined by multilocus sequence analysis". Molecular Ecology. 17 (18): 4092–4106. doi:10.1111/j.1365-294X.2008.03883.x. PMID 19238708. S2CID 38830049.
- Fuhrman J, Davis A (1997). "Widespread Archaea and novel Bacteria from the deep sea as shown by 16S rRNA gene sequences". Marine Ecology Progress Series. 150: 275–285. Bibcode:1997MEPS..150..275F. doi:10.3354/meps150275. ISSN 0171-8630.
- Ivanova, Elena P.; López-Pérez, Mario; Zabalos, Mila; Nguyen, Song Ha; Webb, Hayden K.; Ryan, Jason; Lagutin, Kiril; Vyssotski, Mikhail; Crawford, Russell J.; Rodriguez-Valera, Francisco (2015). "Ecophysiological diversity of a novel member of the genus Alteromonas, and description of Alteromonas mediterranea sp. nov". Antonie van Leeuwenhoek. 107 (1): 119–132. doi:10.1007/s10482-014-0309-y. ISSN 0003-6072. PMID 25326795. S2CID 18595072.
- Ivanova, Elena P.; López-Pérez, Mario; Zabalos, Mila; Nguyen, Song Ha; Webb, Hayden K.; Ryan, Jason; Lagutin, Kiril; Vyssotski, Mikhail; Crawford, Russell J.; Rodriguez-Valera, Francisco (2015-01-01). "Ecophysiological diversity of a novel member of the genus Alteromonas, and description of Alteromonas mediterranea sp. nov". Antonie van Leeuwenhoek. 107 (1): 119–132. doi:10.1007/s10482-014-0309-y. ISSN 1572-9699. PMID 25326795. S2CID 18595072.
- Manck LE, Espinoza JL, Dupont CL, Barbeau KA (April 2020). "Transcriptomic Study of Substrate-Specific Transport Mechanisms for Iron and Carbon in the Marine Copiotroph Alteromonas macleodii". mSystems. 5 (2). doi:10.1128/msystems.00070-20. PMC 7190382. PMID 32345736.
- Goto S, Tada Y, Suzuki K, Yamashita Y (2017). "Production and Reutilization of Fluorescent Dissolved Organic Matter by a Marine Bacterial Strain, Alteromonas macleodii". Frontiers in Microbiology. 8: 507. doi:10.3389/fmicb.2017.00507. PMC 5368175. PMID 28400762.
- Zhang Z, Li Z, Jiao N (June 2014). "Effects of D-amino acids on the EPS production and cell aggregation of Alteromonas macleodii strain JL2069". Current Microbiology. 68 (6): 751–755. doi:10.1007/s00284-014-0520-0. PMID 24526245. S2CID 14922228.
- Rattanaphan P, Mittraparp-Arthorn P, Srinoun K, Vuddhakul V, Tansila N (July 2020). "Indole signaling decreases biofilm formation and related virulence of Listeria monocytogenes". FEMS Microbiology Letters. 367 (14): fnaa116. doi:10.1093/femsle/fnaa116. PMID 32658271.
- Neumann AM, Balmonte JP, Berger M, Giebel HA, Arnosti C, Voget S, et al. (October 2015). "Different utilization of alginate and other algal polysaccharides by marine Alteromonas macleodii ecotypes". Environmental Microbiology. 17 (10): 3857–3868. doi:10.1111/1462-2920.12862. PMID 25847866.
- Mitulla, Maximilian; Dinasquet, Julie; Guillemette, Ryan; Simon, Meinhard; Azam, Farooq; Wietz, Matthias (2016-05-30). "Response of bacterial communities from California coastal waters to alginate particles and an alginolyticAlteromonas macleodiistrain". Environmental Microbiology. 18 (12): 4369–4377. doi:10.1111/1462-2920.13314. ISSN 1462-2912. PMID 27059936.
- Ivars-Martinez E, Martin-Cuadrado AB, D'Auria G, Mira A, Ferriera S, Johnson J, et al. (December 2008). "Comparative genomics of two ecotypes of the marine planktonic copiotroph Alteromonas macleodii suggests alternative lifestyles associated with different kinds of particulate organic matter". The ISME Journal. 2 (12): 1194–1212. doi:10.1038/ismej.2008.74. PMID 18670397. S2CID 520422.
- Roth-Rosenberg D, Aharonovich D, Luzzatto-Knaan T, Vogts A, Zoccarato L, Eigemann F, et al. (August 2020). "Prochlorococcus Cells Rely on Microbial Interactions Rather than on Chlorotic Resting Stages To Survive Long-Term Nutrient Starvation". mBio. 11 (4): 657627. bioRxiv 10.1101/657627. doi:10.1128/mbio.01846-20. PMC 7439483. PMID 32788385.
- Lee MD, Walworth NG, McParland EL, Fu FX, Mincer TJ, Levine NM, et al. (August 2017). "The Trichodesmium consortium: conserved heterotrophic co-occurrence and genomic signatures of potential interactions". The ISME Journal. 11 (8): 1813–1824. doi:10.1038/ismej.2017.49. PMC 5520037. PMID 28440800.
- Fourquez M, Devez A, Schaumann A, Gueneugues A, Jouenne T, Obernosterer I, Blain S (2014-01-27). "Effects of iron limitation on growth and carbon metabolism in oceanic and coastal heterotrophic bacteria". Limnology and Oceanography. 59 (2): 349–360. Bibcode:2014LimOc..59..349F. doi:10.4319/lo.2014.59.2.0349. ISSN 0024-3590. S2CID 85813253.
- Cusick KD, Dale JR, Fitzgerald LA, Little BJ, Biffinger JC (July 2017). "Adaptation to copper stress influences biofilm formation in Alteromonas macleodii". Biofouling. 33 (6): 505–519. doi:10.1080/08927014.2017.1329423. PMID 28604167. S2CID 46739077.
- Chen, Chia-Lung; Maki, James S.; Rittschof, Dan; Teo, Serena Lay-Ming (2013-09-01). "Early marine bacterial biofilm on a copper-based antifouling paint". International Biodeterioration & Biodegradation. 83: 71–76. doi:10.1016/j.ibiod.2013.04.012. ISSN 0964-8305.
- Koch H, Dürwald A, Schweder T, Noriega-Ortega B, Vidal-Melgosa S, Hehemann JH, et al. (January 2019). "Biphasic cellular adaptations and ecological implications of Alteromonas macleodii degrading a mixture of algal polysaccharides". The ISME Journal. 13 (1): 92–103. doi:10.1038/s41396-018-0252-4. PMC 6298977. PMID 30116038.
- Mitulla M, Dinasquet J, Guillemette R, Simon M, Azam F, Wietz M (December 2016). "Response of bacterial communities from California coastal waters to alginate particles and an alginolytic Alteromonas macleodii strain". Environmental Microbiology. 18 (12): 4369–4377. doi:10.1111/1462-2920.13314. PMID 27059936.
- Biller, Steven J.; Coe, Allison; Martin-Cuadrado, Ana-Belen; Chisholm, Sallie W. (2015). "Draft Genome Sequence of Alteromonas macleodii Strain MIT1002, Isolated from an Enrichment Culture of the Marine Cyanobacterium Prochlorococcus". Genome Announcements. 3 (4). doi:10.1128/genomea.00967-15. ISSN 2169-8287. PMC 4551879. PMID 26316635.
- Fadeev, Eduard; De Pascale, Fabio; Vezzi, Alessandro; Hübner, Sariel; Aharonovich, Dikla; Sher, Daniel (2016-03-08). "Why Close a Bacterial Genome? The Plasmid of Alteromonas Macleodii HOT1A3 is a Vector for Inter-Specific Transfer of a Flexible Genomic Island". Frontiers in Microbiology. 7: 248. doi:10.3389/fmicb.2016.00248. ISSN 1664-302X. PMC 4781885. PMID 27014193.
- Kimes NE, López-Pérez M, Ausó E, Ghai R, Rodriguez-Valera F (October 2014). "RNA sequencing provides evidence for functional variability between naturally co-existing Alteromonas macleodii lineages". BMC Genomics. 15 (1): 938. doi:10.1186/1471-2164-15-938. PMC 4223743. PMID 25344729.
- Tada, Yuya; Taniguchi, Akito; Nagao, Ippei; Miki, Takeshi; Uematsu, Mitsuo; Tsuda, Atsushi; Hamasaki, Koji (2011-06-15). "Differing Growth Responses of Major Phylogenetic Groups of Marine Bacteria to Natural Phytoplankton Blooms in the Western North Pacific Ocean". Applied and Environmental Microbiology. 77 (12): 4055–4065. Bibcode:2011ApEnM..77.4055T. doi:10.1128/aem.02952-10. ISSN 0099-2240. PMC 3131633. PMID 21515719.
- Manck, Lauren E.; Park, Jiwoon; Tully, Benjamin J.; Poire, Alfonso M.; Bundy, Randelle M.; Dupont, Christopher L.; Barbeau, Katherine A. (2022). "Petrobactin, a siderophore produced by Alteromonas, mediates community iron acquisition in the global ocean". The ISME Journal. 16 (2): 358–369. doi:10.1038/s41396-021-01065-y. ISSN 1751-7370. PMC 8776838. PMID 34341506.
- Cusick, Kathleen D.; Dale, Jason R.; Little, Brenda J.; Biffinger, Justin C. (2016-12-29). "Draft Genome Sequences of Four Alteromonas macleodii Strains Isolated from Copper Coupons and Grown Long-Term at Elevated Copper Levels". Genome Announcements. 4 (6). doi:10.1128/genomea.01311-16. ISSN 2169-8287. PMC 5122684. PMID 27881542.
- Cusick, Kathleen; Iturbide, Ane; Gautam, Pratima; Price, Amelia; Polson, Shawn; MacDonald, Madolyn; Erill, Ivan (2021-09-28). "Enhanced copper-resistance gene repertoire in Alteromonas macleodii strains isolated from copper-treated marine coatings". PLOS ONE. 16 (9): e0257800. Bibcode:2021PLoSO..1657800C. doi:10.1371/journal.pone.0257800. ISSN 1932-6203. PMC 8478169. PMID 34582496.
- Cusick, Kathleen D.; Polson, Shawn W.; Duran, Gabriel; Hill, Russell T. (2020). "Multiple Megaplasmids Confer Extremely High Levels of Metal Tolerance in Alteromonas Strains". Applied and Environmental Microbiology. 86 (3). Bibcode:2020ApEnM..86E1831C. doi:10.1128/aem.01831-19. ISSN 0099-2240. PMC 6974629. PMID 31757820.
- Colin, M.; Carré, G.; Klingelschmitt, F.; Reffuveille, F.; Gangloff, S. C. (2021-12-15). "Copper alloys to prevent bacterial biofilm formation on touch surfaces". Materials Letters. 305: 130712. doi:10.1016/j.matlet.2021.130712. ISSN 0167-577X.
- Cusick, Kathleen D.; Dale, Jason R.; Fitzgerald, Lisa A.; Little, Brenda J.; Biffinger, Justin C. (2017-07-03). "Adaptation to copper stress influences biofilm formation in Alteromonas macleodii". Biofouling. 33 (6): 505–519. doi:10.1080/08927014.2017.1329423. ISSN 0892-7014. PMID 28604167. S2CID 46739077.
- Gonzaga A, Martin-Cuadrado AB, López-Pérez M, Megumi Mizuno C, García-Heredia I, Kimes NE, et al. (2012-12-01). "Polyclonality of concurrent natural populations of Alteromonas macleodii". Genome Biology and Evolution. 4 (12): 1360–1374. doi:10.1093/gbe/evs112. PMC 3542563. PMID 23212172.
- Garcia-Heredia I, Rodriguez-Valera F, Martin-Cuadrado AB (April 2013). "Novel group of podovirus infecting the marine bacterium 'Alteromonas macleodii.'". Bacteriophage. 3 (2): e24766. doi:10.4161/bact.24766. PMC 3821669. PMID 24228219.
- Feng X, Yan W, Wang A, Ma R, Chen X, Lin TH, et al. (May 2021). "A Novel Broad Host Range Phage Infecting Alteromonas". Viruses. 13 (6): 987. doi:10.3390/v13060987. PMC 8228385. PMID 34073246.
- Bonificio WD, Clarke DR (November 2014). "Bacterial recovery and recycling of tellurium from tellurium-containing compounds by Pseudoalteromonas sp. EPR3". Journal of Applied Microbiology. 117 (5): 1293–1304. doi:10.1111/jam.12629. PMID 25175548. S2CID 18504786.
- Syafitri, E; Prayitno, S B; Ma’ruf, W F; Radjasa, O K (2017). "Genetic diversity of the causative agent of ice-ice disease of the seaweed Kappaphycus alvarezii from Karimunjawa island, Indonesia". IOP Conference Series: Earth and Environmental Science. 55 (1): 012044. Bibcode:2017E&ES...55a2044S. doi:10.1088/1755-1315/55/1/012044. ISSN 1755-1307.
- Cambon-Bonavita, M.-A.; Raguenes, G.; Jean, J.; Vincent, P.; Guezennec, J. (2002). "A novel polymer produced by a bacterium isolated from a deep-sea hydrothermal vent polychaete annelid". Journal of Applied Microbiology. 93 (2): 310–315. doi:10.1046/j.1365-2672.2002.01689.x. ISSN 1364-5072. PMID 12147080. S2CID 23606744.
- Le Costaouëc T, Cérantola S, Ropartz D, Ratiskol J, Sinquin C, Colliec-Jouault S, Boisset C (September 2012). "Structural data on a bacterial exopolysaccharide produced by a deep-sea Alteromonas macleodii strain". Carbohydrate Polymers. 90 (1): 49–59. doi:10.1016/j.carbpol.2012.04.059. PMID 24751009.
Further reading
- "Species: Alteromonas macleodii". List of Prokaryotic names with Standing in Nomenclature LPSN. Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures GmbH. Retrieved 2022-04-04.
External links
- Podstawka, Adam (2021-12-21). "Alteromonas macleodii 107 - Type strain - DSM 6062, ATCC 27126, CCUG 16128, CIP 103198, JCM 20772, LMG 2843, NBRC 102226, IAM 12920, NCIMB 1963". BacDive. BacDiveID:444. Retrieved 2022-04-04.