Anti-miRNA oligonucleotides
Anti-miRNA oligonucleotides (also known as AMOs) have many uses in cellular mechanics. These synthetically designed molecules are used to neutralize microRNA (miRNA) function in cells for desired responses. miRNA are complementary sequences (≈22 bp) to mRNA that are involved in the cleavage of RNA or the suppression of the translation.[1] By controlling the miRNA that regulate mRNAs in cells, AMOs can be used as further regulation as well as for therapeutic treatment for certain cellular disorders. This regulation can occur through a steric blocking mechanism as well as hybridization to miRNA.[2] These interactions, within the body between miRNA and AMOs, can be for therapeutics in disorders in which over/under expression occurs or aberrations in miRNA lead to coding issues. Some of the miRNA linked disorders that are encountered in the humans include cancers, muscular diseases, autoimmune disorders, and viruses. In order to determine the functionality of certain AMOs, the AMO/miRNA binding expression (transcript concentration) must be measured against the expressions of the isolated miRNA. The direct detection of differing levels of genetic expression allow the relationship between AMOs and miRNAs to be shown. This can be detected through luciferase activity (bioluminescence in response to targeted enzymatic activity). Understanding the miRNA sequences involved in these diseases can allow us to use anti miRNA Oligonucleotides to disrupt pathways that lead to the under/over expression of proteins of cells that can cause symptoms for these diseases.
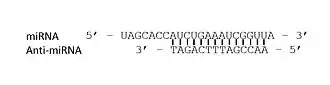
Synthesis
During anti-miRNA oligonucleotide design, necessary modifications to optimize binding affinity, improve nuclease resistance, and in vivo delivery must be considered.[3] There have been several generations of designs with attempts to develop AMOs with high binding affinity as well as high specificity. The first generation utilized 2’-O-Methyl RNA nucleotides with phosphorothioate internucleotide linkages positioned at both ends to prevent exonuclease attack. A recent study discovered a compound, N,N-diethyl-4-(4-nitronaphthalen-1-ylazo)-phenylamine (ZEN), that improved binding affinity and blocked exonuclease degradation.[4] This method was combined with the first generation design to create a new generation ZEN-AMO with an improved effectiveness.
Various components of AMOs can be manipulated to affect the binding affinity and potency of the AMO. The 2’-sugar of the AMOs can be modified to be substituted with fluorine and various methyl groups, almost all with an increase in binding affinity. However, some of these modified 2’-sugar AMOs led to negative effects on cell growth. Modifying the 5'-3' phosphodiester backbone linkage to a phosphorothiorate (P-S) backbone linkage was also shown to have an effect on target affinity. Using the P-S mutation was shown to decrease the Tm of the oligonucleotide, which leads to a lower target affinity. A final requirement for AMOs is mismatch specificity and length restrictions. Due to miRNAs in the same families sharing “seed” (shared) sequences and differ by only a couple of additional nucleotides; one AMO can potentially target multiple miRNA sequences. However, studies have suggested that this is difficult due to the loss of activity with single nucleotide mismatches. Greater than three mismatches demonstrates complete loss of activity. Changes in the length of AMOs were tolerated far better, with changes of one nucleotide and two nucleotides resulting in little loss of activity and three or more in total loss of activity. Truncating a single nucleotide from the 3’ end resulted in a slight improvement of AMO activity.[5]
Delivery and Detection

Delivery of AMOs requires in vitro transfection into target cells. Presently there are difficulties with conventional methods of transfection that result in low delivery efficiency. In order to increase the effectiveness of AMO delivery, a 2011 paper proposed using functionalized gold nanoparticles. The gold nanoparticles increase delivery efficiency by conjugating with a cargo DNA that anneals to the AMO using complementarity. The cargo DNA is attached to the surface of the nanoparticle.[6] Because many variations of DNA and RNA are unstable in in vivo conditions, carriers, such as nanoparticles, are necessary to protect from degeneration by nucleases. These nanoparticles are useful in order to facilitate uptake into the cell, and transfer the genetic information to the nucleus.[7] Another in vivo method for delivery supported by results in mice is the injection of AMOs intravenously. Tail vein injection of AMOs in the mice were shown to be effective. In order for this system to be useful, the AMOs were conjugated with cholesterol for increased uptake into the cell through the membrane and were chemically modified by 2′-OMe phosphoramidites to prevent degradation of the AMOs.[8]
To detect the presence and functionality of AMOs, researchers can observe the relative activity of the target enzyme or protein of the miRNA. This method was used in a study of single AMOs targeting multiple miRNAs, where relative luciferase activity in HEK293 cells was monitored. To determine relative Luciferase activity levels, a control with no miRNA present was included. The presence of functional AMOs with the inhibiting miRNA would result in an increase in Luciferase activity due to the inactivation of the miRNA suppressing the enzyme's activity.[9]
Disorders/Therapeutics
Many human disorders have been found to have some alterations in expression or aberrations involving miRNA. It has been found that miRNA have been involved in many key regulation pathways that are suspected to be related to cancer, viral genes, and metabolic pathways,[10] as well as muscular disorders (specifically cardiovascularly related).[11] By targeting cells affected with improper miRNA expression, the normal balance of the expression can be restored by using AMOs. By minimizing overexpression and increasing underexpression with AMOs, some of these genetic disorders can be potentially bypassed or at least have their symptoms minimized. This is done by hybridization of the AMOs to miRNA sequences that are involved in the expression of specific genes. The issue is finding a way for the AMOs to successfully perform their function in concentrations that are sufficient for success, while at the same time being low enough to avoid toxicity of the vector and AMOs themselves.
Cancer
All cancers are mutations in the genomes that cause abnormal cell growth. Determining factors that contribute to or regulate this excessive growth can potentially lead to preventative, therapeutic treatments of cancer. For example, chronic lymphocytic leukemias illustrates a region of miRNAs (mir-15 and mir-16) are missing from the genome in the expression of this cancer. While in other cancers, such as Burkitt's lymphoma, expression of miRNA sequences are amplified.[10] This leads to the suggestion that many miRNA have regulatory sequences involved in cancer. If those were to be better regulated, potentially through AMOs, perhaps the onset and progression of cancer could be regulated.
Following a study of 540 tumor samples of various cancer types, it was discovered that 15 miRNAs were upregulated and 12 were downregulated.[12] From the study, it was concluded that these miRNA sequences had an effect on cell growth and apoptosis in the cell. AMOs play into the equation as this regulatory factor for the miRNAs involved in cancer. If bound to a single affected miRNA site, the effect appears to be minimal. However, by creating sequences of anti-miRNA Oligonucleotides to bind to all of these implicit miRNAs, there was increased cell death within the cancer cells.[11] One study involving antagomirs, a different variation of anti-miRNA oligonucleotides, focused on reducing induced tumors in mice. After 2 weeks of treatment, tumor growth was inhibited and regression was shown in 30% of cases.[13] This illustrates that AMOs can be used to successfully inhibit cancers through miRNAs. This inhibition is caused by a direct silencing interaction of the miRNAs that in turn bind on the mRNA sequences that create proteins in cancer cells, as well as increased control of cellular processes of cancer.
Muscular Development
In the development of tissues in embryos, miRNA can have a role in the upregulation or downregulation of specific muscular development. miRNA-1 plays a role in muscle differentiation between cardiac and skeletal muscle precursor cells.[14] In development, if levels of precursor cells are not properly regulated, it can result in muscular hypoplasia. By creating AMOs for these known miRNAs involved in muscle generation, it is possible to track a miRNA's specific mechanisms throughout the process of muscle generation by essentially using the created AMO to turn off the miRNA. This halts the production of myogenin(the transcription factor involved in myogenesis). By then measuring the changes in myogenin compared to standard, non-inhibited myogenesis, a miRNA's function can be determined as either upregulating or downregulating the synthesis of myogenin.[15] By further understanding how certain miRNA sequences control the development of muscle, AMOs can be utilized to promote normal production levels of myogenin in organisms that have been detected to contain genetic errors involving myogenesis.
AMOs can also be used to prevent apoptosis, or organ hypoplasia, of the heart in the presence of high concentrations of hydrogen peroxide. Hydrogen peroxide can induce apoptosis through oxidative stress. This is because oxidative stress caused by H
2O
2 induces increased activity of miRNA-1. This increased miRNA-1 activity represses the activity of Bcl-2, inducing apoptosis. However, by creating and introducing an AMO for miRNA-1 in an environment of oxidative stress, the response to H
2O
2 is reduced, creating a resistance to oxidative stress in the heart. Because of this, the amount of hydrogen peroxide-induced apoptosis of cardiomyocytes is reduced in heart disease.[16] Due to the reduction of cardiomyocyte death in conditions of oxidative stress by the anti-miRNA-1 Oligonucleotide, miRNA regulation can allow us to more deeply understand the development of the heart, as well as the survival of heart muscle in low oxygen conditions.
Autoimmune Response and Disorders
Autoimmune disorders are when the body has an immune response to itself, causing an inflammatory reaction to occur within the body. Because autoimmune disorders involve abnormalities in the immune system cells (i.e., B-cells, T-cells). It can be inferred that miRNA are strongly expressed in regions of the body that have to do with the maturation of these T and B lymphocytes, such as in the spleen and lymph nodes.[17] Abnormalities in the miRNA or the function of the miRNA in the post-transcriptional process can result in an increased sensitivity of the lymphocytes. Due to increased sensitivity, these lymphocytes can now target antigens that it could not previously bind, which can allow for these lymphocytes to attack itself, if these antigens happen to naturally occur in cells in the body.[18]
One instance of this is Rheumatoid Arthritis, in which the body breaks down its own joints. The breakdown is caused by the overexpression of specific miRNA clusters. These clusters causes an increase in synovial fibroblasts. Due to this increased fibroblast amount, certain proteases' concentrations are increased which cause the breakdown of cartilage in joints.[17] By targeting the miRNA clusters responsible for expression of the disease, inflammation caused by this disorder can be reduced when AMOs are added to afflicted areas.
Systemic lupus erythematosus causes long term organ damage to the body. It propagates due to environmental and genetic factors. By targeting microRNA (miR-184, miR-196a, miR-198, and miR-21)[19] that are down-regulated in SLE with AMOs in the affected organs, the normal expression of these genes can be restored.
Viral Studies
It is believed that cellular miRNAs inhibit viral gene expression. In a study of HIV-1, anti-miRNA inhibitors were used to deactivate two miRNAs that inhibit viral gene expression, has-miR-29a and 29b. It was shown that viral gene expression increased after the introduction of anti-miRNAs targeting has-miR-29a and 29b. This demonstrated miRNA inhibitors were able to directly target and reverse the inhibitory effect of has-miR-29a and 29b in the HIV-1 virus.[20] By creating an AMO, certain genomic sequences of HIV were able to be studied more in depth. A further understanding of the way the genome of certain viruses work can allow scientists to create preventative measures against these viruses.
The mechanism for anti-miRNA regulation in the case of the Epstein-Barr virus (EBV) differs slightly than other viral cases such as HIV-1. EBV is a herpesvirus related to various cancers that has the ability to express miRNAs, unlike many other viruses that affect humans. Unlike other studies that utilize anti-miRNAs as a knockdown tool to demonstrate the effects of miRNAs, researchers of EBV used them to inhibit the miRNAs produced by the virus. MiR-BART5, a miRNA of EBV, regulates the protein: p53 Up-regulated Modulator of Apoptosis (PUMA). When the viral mir-BART5 was depleted using its anti-miRNA, anti-miR-BART5, cell apoptosis was triggered and resulted in disease control, killing the cells that are identified as infected.[21]
Another peculiar case of host-viral interaction mediated by a microRNA occurs with the hepatitis C virus (HCV). HCV, which causes acute infection of the liver, often going undetected and progressing to chronic, uses the human miRNA miR-122 to recruit Argonaute2 proteins to the uncapped 5' end of its RNA genome, thereby masking it from the cellular antiviral response and stabilizing it. This interaction has led to the development of AMOs that target miR-122 in an effort to clear the virus from the hepatic cells.[22] The most advanced of these compounds is miravirsen, a locked nucleic acid-DNA mixmer, [23] currently undergoing clinical trials.[24] An interesting aspect of miravirsen is its reported ability to inhibit not just the mature miR-122, but to also invade the stem-loop structures in the microRNA precursor molecules, disrupting the biogenesis of miR-122 in biochemical assays and cell culture.[25]
References
- Cox, David L. Nelson, Michael M. (2008). Lehninger principles of biochemistry (5th ed.). New York: W.H. Freeman. p. 1045. ISBN 978-0-7167-7108-1.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Lennox, K A; Behlke, M A (14 July 2011). "Chemical modification and design of anti-miRNA oligonucleotides". Gene Therapy. 18 (12): 1111–1120. doi:10.1038/gt.2011.100. PMID 21753793.
- Stenvang, Jan; Petri, Andreas; Lindow, Morten; Obad, Susanna; Kauppinen, Sakari (2012). "Inhibition of microRNA function by antimiR oligonucleotides". Silence. 3 (1): 1. doi:10.1186/1758-907X-3-1. PMC 3306207. PMID 22230293.
- Lennox, Kim A; Owczarzy, Richard; Thomas, Derek M; Walder, Joseph A; Behlke, Mark A (2013). "Improved Performance of Anti-miRNA Oligonucleotides Using a Novel Non-Nucleotide Modifier". Molecular Therapy: Nucleic Acids. 2 (8): e117. doi:10.1038/mtna.2013.46. PMC 3759741. PMID 23982190.
- Davis, S.; Lollo, B; Freier, S; Esau, C (28 April 2006). "Improved targeting of miRNA with antisense oligonucleotides". Nucleic Acids Research. 34 (8): 2294–2304. doi:10.1093/nar/gkl183. PMC 1459537. PMID 16690972.
- Kim, Jae-Hong; Yeom, Ji-Hyun; Ko, Jeong-Jae; Han, Min Su; Lee, Kangseok; Na, Soon-Young; Bae, Jeehyeon (September 2011). "Effective delivery of anti-miRNA DNA oligonucleotides by functionalized gold nanoparticles". Journal of Biotechnology. 155 (3): 287–292. doi:10.1016/j.jbiotec.2011.07.014. PMID 21807040.
- Neu, Michael; Fischer, Dagmar; Kissel, Thomas (August 2005). "Recent advances in rational gene transfer vector design based on poly(ethylene imine) and its derivatives". The Journal of Gene Medicine. 7 (8): 992–1009. doi:10.1002/jgm.773. PMID 15920783. S2CID 25022652.
- Krützfeldt, Jan; Rajewsky, Nikolaus; Braich, Ravi; Rajeev, Kallanthottathil G.; Tuschl, Thomas; Manoharan, Muthiah; Stoffel, Markus (30 October 2005). "Silencing of microRNAs in vivo with 'antagomirs'". Nature. 438 (7068): 685–689. Bibcode:2005Natur.438..685K. doi:10.1038/nature04303. PMID 16258535. S2CID 4414240.
- Lu, Y.; Xiao, J.; Lin, H.; Bai, Y.; Luo, X.; Wang, Z.; Yang, B. (9 January 2009). "A single anti-microRNA antisense oligodeoxyribonucleotide (AMO) targeting multiple microRNAs offers an improved approach for microRNA interference". Nucleic Acids Research. 37 (3): e24. doi:10.1093/nar/gkn1053. PMC 2647303. PMID 19136465.
- Weiler, J; Hunziker, J; Hall, J (29 September 2005). "Anti-miRNA oligonucleotides (AMOs): ammunition to target miRNAs implicated in human disease?". Gene Therapy. 13 (6): 496–502. doi:10.1038/sj.gt.3302654. PMID 16195701.
- Wang, Zhiguo (28 August 2010). "The Concept of Multiple-Target Anti-miRNA Antisense Oligonucleotide Technology". MicroRNA and Cancer. Methods in Molecular Biology. Vol. 676. pp. 51–57. doi:10.1007/978-1-60761-863-8_4. ISBN 978-1-60761-862-1. PMID 20931389.
- Volinia, S.; Calin, G. A.; Liu, C.-G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; Prueitt, R. L.; Yanaihara, N.; Lanza, G.; Scarpa, A.; Vecchione, A.; Negrini, M.; Harris, C. C.; Croce, C. M. (3 February 2006). "A microRNA expression signature of human solid tumors defines cancer gene targets". Proceedings of the National Academy of Sciences. 103 (7): 2257–2261. doi:10.1073/pnas.0510565103. PMC 1413718. PMID 16461460.
- Negrini, Massimo; Nicoloso, Milena S; Calin, George A (2009). "MicroRNAs and cancer—new paradigms in molecular oncology". Current Opinion in Cell Biology. 21 (3): 470–479. doi:10.1016/j.ceb.2009.03.002. PMID 19411171.
- Zhao, Yong; Samal, Eva; Srivastava, Deepak (14 July 2005). "Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis". Nature. 436 (7048): 214–220. Bibcode:2005Natur.436..214Z. doi:10.1038/nature03817. PMID 15951802. S2CID 4340449.
- Chen, Jian-Fu; Mandel, Elizabeth M; Thomson, J Michael; Wu, Qiulian; Callis, Thomas E; Hammond, Scott M; Conlon, Frank L; Wang, Da-Zhi (25 December 2005). "The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation". Nature Genetics. 38 (2): 228–233. doi:10.1038/ng1725. PMC 2538576. PMID 16380711.
- Tang, Yehua; Zheng, Jiaoyang; Sun, Yan; Wu, Zonggui; Liu, Zhimin; Huang, Gaozhong (2009). "MicroRNA-1 Regulates Cardiomyocyte Apoptosis by Targeting Bcl-2". International Heart Journal. 50 (3): 377–387. doi:10.1536/ihj.50.377. PMID 19506341.
- Tili, Esmerina; Michaille, Jean-Jacques; Costinean, Stefan; Croce, Carlo M (26 August 2008). "MicroRNAs, the immune system and rheumatic disease". Nature Clinical Practice Rheumatology. 4 (10): 534–541. doi:10.1038/ncprheum0885. PMID 18728632. S2CID 25869468.
- "Immune response". Medline Plus. National Library of Medicine. Retrieved 1 November 2014.
- Kaucsár, Tamás; Rácz, Zsuzsanna; Hamar, Péter (30 November 2010). "Post-transcriptional gene-expression regulation by micro RNA (miRNA) network in renal disease☆". Advanced Drug Delivery Reviews. 62 (14): 1390–1401. doi:10.1016/j.addr.2010.10.003. PMID 20940025.
- Ahluwalia, Jasmine K; Khan, Sohrab; Soni, Kartik; Rawat, Pratima; Gupta, Ankit; Hariharan, Manoj; Scaria, Vinod; Lalwani, Mukesh; Pillai, Beena; Mitra, Debashis; Brahmachari, Samir K (2008). "Human cellular microRNA hsa-miR-29a interferes with viral nef protein expression and HIV-1 replication". Retrovirology. 5 (1): 117. doi:10.1186/1742-4690-5-117. PMC 2635386. PMID 19102781.
- Choy, E. Y.-W.; Siu, K.-L.; Kok, K.-H.; Lung, R. W.-M.; Tsang, C. M.; To, K.-F.; Kwong, D. L.-W.; Tsao, S. W.; Jin, D.-Y. (6 October 2008). "An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival" (PDF). Journal of Experimental Medicine. 205 (11): 2551–2560. doi:10.1084/jem.20072581. PMC 2571930. PMID 18838543.
- Sarnow, P; Sagan, SM (2016). "Unraveling the Mysterious Interactions Between Hepatitis C Virus RNA and Liver-Specific MicroRNA-122". Annual Review of Virology. 3 (1): 309–332. doi:10.1146/annurev-virology-110615-042409. PMID 27578438.
- Elmén J, Lindow M, Schütz S, Lawrence M, Petri A, Obad S, Lindholm M, Hedtjärn M, Hansen HF, Berger U, Gullans S, Kearney P, Sarnow P, Straarup EM, Kauppinen S (2008). "LNA-mediated microRNA silencing in non-human primates". Nature. 452 (7189): 896–9. Bibcode:2008Natur.452..896E. doi:10.1038/nature06783. PMID 18368051. S2CID 4308734.
- Janssen, Harry L.A.; Reesink, Hendrik W.; Lawitz, Eric J.; Zeuzem, Stefan; Rodriguez-Torres, Maribel; Patel, Keyur; Van Der Meer, Adriaan J.; Patick, Amy K.; Chen, Alice; Zhou, Yi; Persson, Robert; King, Barney D.; Kauppinen, Sakari; Levin, Arthur A.; Hodges, Michael R. (2013). "Treatment of HCV Infection by Targeting MicroRNA". New England Journal of Medicine. 368 (18): 1685–94. doi:10.1056/NEJMoa1209026. PMID 23534542.
- Gebert, Luca F. R.; Rebhan, Mario A. E.; Crivelli, Silvia E. M.; Denzler, Rémy; Stoffel, Markus; Hall, Jonathan (7 January 2014). "Miravirsen (SPC3649) can inhibit the biogenesis of miR-122". Nucleic Acids Research. 42 (1): 609–621. doi:10.1093/nar/gkt852. PMC 3874169. PMID 24068553.