Archer City Formation
The Archer City Formation is a geological formation in north-central Texas, preserving fossils from the Asselian and early Sakmarian stages of the Permian period. It is the earliest component of the Texas red beds, introducing an tropical ecosystem which will persist in the area through the rest of the Early Permian. The Archer City Formation is preceded by the cool Carboniferous swamp sediments of the Markley Formation, and succeeded by the equally fossiliferous red beds of the Nocona Formation. The Archer City Formation was not named as a unique geological unit until the late 1980s. Older studies generally labelled its outcrops as the Moran or Putnam formations, which are age-equivalent marine units to the southwest.
| Archer City Formation | |
|---|---|
| Stratigraphic range: Early Permian, Asselian–early Sakmarian | |
| Type | Formation |
| Unit of | Bowie Group |
| Underlies | Nocona Formation |
| Overlies | Markley Formation |
| Lithology | |
| Primary | mudstone |
| Other | sandstone, siltstone |
| Location | |
| Region | |
| Country | |
| Type section | |
| Named for | Archer City, Texas |
| Named by | Hentz & Brown, 1987 |
 | |
The sediments of the Archer City Formation reconstruct a coastal floodplain crossed by small meandering rivers and isolated ponds.[1][2] The climate was hot and monsoonal, preserving a variety of paleosols and plant assemblages ranging from dry upland forests to moist riverside galleries. "Seed ferns" (Sphenopteris, Autunia, Odontopteris, etc.) and early conifers (Walchia, etc.) occupied drier soils, while marattialean ferns (Pecopteris, Asterotheca, etc.) and horsetails (Sphenophyllum, Annularia, Calamites, etc.) were prominent in more humid areas.
The Archer City Formation preserves a diverse fauna of fossil vertebrates in pond bonebeds across Archer and Clay counties. Many fossils are rare fragments of species which are better-preserved in younger parts of the red beds. Nevertheless, complete skeletons and important type specimens are also known from the formation. Some notable synapsids include Apsisaurus witteri, Edaphosaurus boanerges, Ophiacodon uniformis, and Dimetrodon milleri. Early reptiles such as Protorothyris archeri and Romeria spp. make up a small component of the terrestrial fauna. Amphibians are common in every niche, with examples such as Phonerpeton pricei, Neldasaurus wrightei, Edops craigi, Eryops megacephalus, and Pantylus cordatus, just to name a few. The rivers were roamed by Orthacanthus texensis (a xenacanth "shark") and Ectosteorhachis nitidus (a megalichthyid fish).
History and stratigraphy
Fossil bonebeds in the vicinity of Archer City were first investigated in detail by Alfred Romer and Llewellyn Price, starting in the late 1920s.[3][4][5][6] At the time, these bonebeds were assigned to the Pueblo, Moran, or Putnam formations of the Cisco Group. The Archer City Formation was first named in a 1987 geologic map seeking to update the stratigraphy of North Texas.[7] The formation and its stratigraphic placement were formalized in text a year later.[8]
The formation outcrops in an arc from Montague County in the east, through Clay, Archer, and Young counties, as far as Throckmorton County in the west. It is most broadly exposed in Clay and Archer counties, including under its namesake of Archer City.[7][8][9]
The Archer City Formation is a unit of the Bowie Group, lying above the Markley Formation and below the Nocona Formation of the Wichita Group.[7][8][1] Some publications have labelled the Nocona Formation as the "Upper Archer City Formation", thus considering the Archer City Formation to be a unit of the Wichita Group.[10] This proposal is not widely agreed upon.[11][9] The Cisco Group and its constituent formations have been restricted to marine strata further south and west. Of these strata, the Archer City Formation is roughly equivalent in time to the uppermost Pueblo Formation, the Moran Formation, the Sedwick Formation, and the Santa Anna Branch shale (= Putnam Formation), from oldest to youngest.[7][8][2]
Age
Based on correlations with coastal strata further west, the Carboniferous-Permian boundary is positioned near the top of the Markley Formation. This suggests that the Archer City Formation occupies most of the Asselian (299–293 Ma), the first global stage of the Permian Period.[11] The plant fossils of the Archer City Formation also resemble those of Asselian Europe.[9] The upper part of the Archer City Formation may extend into the succeeding Sakmarian stage (293–290 Ma). The Santa Anna Branch Shale and overlying Coleman Junction Limestone both preserve fossils of Sweetognathus merrilli,[12] a conodont index fossil of the early Sakmarian. The Asselian and Sakmarian global stages are equivalent to the early-middle parts of the Wolfcampian regional stage.[13]
Paleoenvironment
As with the rest of the Texas red beds, the Archer City Formation was deposited on a coastal plain between an inland marine basin in the west and patches of steadily eroding mountains in the north and east. Some of the mountains are still standing in the present day, such as the Wichita Mountains and Arbuckle Mountains, while others have been fully eroded or buried, such as the Texan portion of the Ouachita Mountains. The marine basin is formally known as the Midland Basin, a major component of the Permian Basin oil field.[8][2] Rising sea levels through the Early Permian allowed the coastline to gradually transgress northward and eastward. The major bonebed sites of the Archer City Formation were probably 40–60 km (25–37 miles) away from the coast.[2]
During the Permian, the area was near the equator, with a latitude of 0° to 5° N.[2] Temperature estimates can be inferred for the Archer City Formation via δ18O records for hematite and phyllosilicate minerals in paleosols. These data points suggest soil temperatures of 31–35 °C (88–95 °F), significantly warmer than the cool and wet conditions of the Markley Formation.[11] This fits with climate evidence from rapid sedimentary changes between the formations, as well as an increasingly patchy distribution of high-humidity flora.[2][13][11] The Archer City Formation would have had a hot monsoonal[2] climate, with moist or swampy areas shrinking to the vicinity of riverbanks and ponds during the dry season.[13]
Geology
In terms of their lithologies, the Archer City and Nocona formations are nearly indistinguishable,[10] with equivalent sets of fluvial and floodplain sediments.[1] The vast majority of each formation is homogeneous red mudstone with small calcareous nodules. Fluvial sandstone beds[1] and paleosols[2] are also common. Fossils and other organic debris are concentrated into rare greyish claystone bonebeds, which correspond to perennial ponds. Most of the pond bonebeds are too small to continuously support an ecosystem of large aquatic vertebrates, suggesting that the carcasses may have been washed in from larger bodies of water.[1]
Sandstone beds
By volume, the most prevalent sandstone beds are upwards-fining stacks of point bar, scroll-bar, and coarser channel fill deposits. These are all hallmarks of small meandering rivers, with channels only a few meters wide.[1] Other common sandstone features include subdued, fine-grained crevasse splay deposits. The crevasse splays are often associated with plant fossils which have been remineralized by copper minerals.[1]
Some sandstone beds are isolated straight channel fills, without corresponding riverbank deposits.[1] These straight beds may reach 30 meters in width (though most are much narrower) and several kilometers in length. Coarser beds preserve sinuous megaripples while finer channels develop small climbing ripples towards the top. Straight channels would have formed during high-energy erosion events, but the climate was probably too moist for frequent flash floods.[1]
Paleosols
Four different types of paleosol (labelled types E through H) have been identified in the Archer City Formation.[2][13] Three (F, G, and H) first appear in this formation, while type E paleosols are retained from the underlying Markley Formation. All four continue to persist through the Texas Lower Permian up until an aridification event in the mid-Clear Fork Group. The paleosols are generally dark red or brown, with only intermittent gleying in the type E variety. The Archer City Formation mostly lacks the high-humidity paleosol varieties of the Markley Formation (types A through D).[2][13]
Type E paleosols are silty or fine sandy soils with persistent mottling, hematite nodules, and few distinct horizons. They are a type of entisol, young soils corresponding to isolated, frequently disturbed areas with a high water table, such as riverbanks.[2][13] Type F paleosols are alfisols, patterned with deep root casts and numerous horizons, including intermediate layers with calcareous nodules and smectite clay. They develop in stable forested uplands with a low water table and a semi-arid to subhumid climate (~70 cm of annual precipitation).[2][13] Type G paleosols are similar, with sandy vertical cracks and abundant calcareous nodules. They qualify as vertisols, inundated and dried at a seasonal frequency along the margins of a floodplain.[2][13] Type H paleosols have an even greater proportion of calcareous minerals, forming blocky crusts interspersed among chlorite- and mica-rich clay. They could be termed aridisols or (more broadly) inceptisols, deposited in dry upland areas free from the influence of groundwater.[2][13]
Paleobiota
Color key
|
Notes Uncertain or tentative taxa are in small text; |
Synapsids
| Synapsids of the Archer City Formation | |||
|---|---|---|---|
| Species | Material | Notes | Images |
| Apsisaurus witteri[14] | A partial skeleton | A varanopid,[15] originally identified as a diapsid reptile.[14] | |
| Dimetrodon milleri[16] | Nearly complete skeleton[16][5][1][17] | A small sail-backed sphenacodontid |  |
| Edaphosaurus boanerges[5] | Isolated neural spines and at least one partial skeleton[1][17] | A common medium-sized edaphosaurid, often confused with Edaphosaurus cruciger.[5] |  |
| Lupeosaurus kayi[16] | Limb and hip bones and vertebrae[16][5][17] | A small basal edaphosaurid[18] | |
| Ophiacodon retroversus | Rare fragments[5] | A large ophiacodontid, also described under the names Theropleura and Diopeus. |  |
| Ophiacodon uniformis | A partial skeleton with a well-preserved skull[1][5] | A medium-sized ophiacodontid, also described under the name Poliosaurus.[5] |  |
| Stereophallodon ciscoensis[16] | Skull fragments and vertebrae[5][17][10] | A large ophiacodontid | 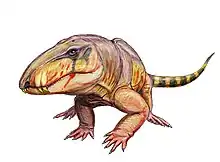 |
Reptiles
| Reptiles of the Archer City Formation | |||
|---|---|---|---|
| Species | Material | Notes | Images |
| Bolosaurus striatus | A jaw[1][17] | A bolosaurid parareptile |  |
| Protorothyris archeri[19] | Five skulls with associated postcranial material[20][17] | A protorothyridid eureptile |  |
| Romeria prima[20] | A skull[20] | A captorhinid eureptile | |
| Romeria texana[19] | A partial skull and partial skeleton[1][20] | A captorhinid eureptile | |
Temnospondyls
| Temnospondyls of the Archer City Formation | |||
|---|---|---|---|
| Species | Material | Notes | Images |
| Acheloma cumminsi | Partial skull and postcrania[21][1] | A trematopid |  |
| Aspidosaurus sp. | Neural spine armor fragments[22][23] | A dissorophid | |
| Broiliellus brevis[22] | Two skulls, one with a partial skeleton[22] | A dissorophid |  |
| Brevidorsum profundum[22] | Partial skull and postcrania[22][17][23] | A dissorophid | |
| Diploseira angusta[24] | A partial skeleton[22] | A dissorophid, formerly considered a species of Dissorophus.[22] | |
| Edops craigi | Several skulls and postcrania[25][17][10] | An edopoid |  |
| Eryops megacephalus | Jaws and other fragments[1][17][10][26] | A common eryopid | 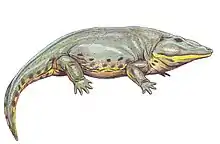 |
| Neldasaurus wrightae[27] | Four partial skulls and postcania[27][17][10][28] | A trimerorhachid dvinosaur | 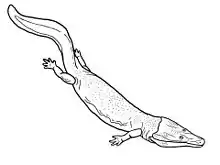 |
| "Parioxys" bolli[29] | A partial skeleton[29][17] | A temnospondyl of uncertain affinities. Parioxys ferricolus, the more complete type species of Parioxys, has variably been interpreted as an eryopoid or a dissorophid."Parioxys" bolli is based on more fragmentary fossils and may belong to an unrelated genus, perhaps a trematopid related to Ecolsonia cutlerensis.[30] | |
| Pasawioops cf. mayi | A skull[31] | A micropholid amphibamiform | |
| Phonerpeton pricei[32] | At least seven skulls with associated postcranial fragments[32] | A trematopid, formerly considered a species of Acheloma.[21] |  |
| Reiszerpeton renascentis | A skull[31] | A dissorophid | |
| Tersomius texensis | A skull[17][31] | A micropholid amphibamiform | |
| Trimerorhachis sp. | A trimerorhachid dvinosaur. Reports from the Archer City Formation[17][10] are unsubstantiated, though it is known from time-equivalent strata in New Mexico.[33] | ||
| Zatrachys serratus | Skull fragments[1][17][10] | A zatracheid |  |
Other amphibians
| Non-temnospondyl amphibians of the Archer City Formation | |||
|---|---|---|---|
| Species | Material | Notes | Images |
| Archeria crassidisca | Jaws and vertebrae[34][17][1] | An archeriid embolomere, often conflated with the more fragmentary genus Cricotus.[34] |  |
| Diadectes sideropelicus | Jaws and vertebrae[3][4][17][1] | A diadectomorph | .jpg.webp) |
| Diplocaulus sp. | Vertebrae[4][17] | A diplocaulid nectridean |  |
| Pantylus cordatus | Multiple skeletons in varying states of completeness[35][17][1] | A pantylid microsaur | 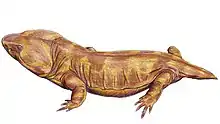 |
| Sauropleura bairdi | Partial skeleton[36] | A urocordylid nectridean |  |
| Seymouria baylorensis | Vertebrae[1] | A seymouriamorph | .jpg.webp) |
Fish
Scales of indeterminate palaeoniscoids, spines of Xenacanthus, Hybodus, Ctenacanthus, and "Anodontacanthus americanus" (= Platyacanthus),[37] and teeth of Barbclabornia luederensis, Helodus, Gnathorhiza, petalodonts, and cladodonts have also been reported from bonebeds of the Archer City Formation.[38][1][39]
| Fish of the Archer City Formation | |||
|---|---|---|---|
| Species | Material | Notes | Images |
| Ectosteorhachis nitidus | Scales, jaws, and a skull[40][1][39] | A megalichthyid tetrapodomorph |  |
| Orthacanthus compressus | Teeth[39] | A xenacanth "shark" | |
| Orthacanthus texensis | Teeth, fin spines, cartilage fragments, and a skull[1][39] | A xenacanth "shark" |  |
| Sagenodus sp. | Tooth plates[1] | A lungfish | |
Plants
Plant fossils in the Archer City Formation are concentrated at two sites in Clay County: Kola Switch and Sanzenbacher Ranch.[9][41] Both sites were discovered in 1940–1941 by Adolph H. Witte, a local geologist working for the WPA. Though Witte's original collections have been lost, he sampled the sites further in 1961, along with USGS geologists Sergius H. Mamay and Arthur D. Watt. On behalf of the USNM, Mamay led another expedition in 1990–1991.[9][41]
The floral diversity of the Archer City Formation strongly overlaps with other Asselian-Sakmarian formations in the southwestern United States. These include the Bursum and Abo formations of New Mexico and the Neal Ranch Formation of West Texas. There are some notable differences, such as a lack of marattialean ferns in the Bursum Formation and a greater variety of swamp foliage in the Neal Ranch Formation. Many of the broader groups prevalent in these formations are equally abundant in the Rotliegend of Europe, even if only a few specific species (such as Autunia conferta) are common in both regions. Conversely, there is little similarity with swampy coal-bearing areas such as the Dunkard Group of the Appalachian Basin. This is most likely a consequence of local environmental factors rather than a turnover of plant diversity at the Permo-Carboniferous boundary.[41][9]
Kola Switch
The Kola Switch site is divided into three different layers in a 1.4 meter (4.6 feet) thick interval, each with a different set of fossils. The lower bed is pale upwards-fining siltstone, the middle bed is dark shale with carbonaceous impressions, and the upper bed is greenish claystone.[9]
The seed fern Sphenopteris germanica is the most abundant plant fossil in the lower bed. The conifer Walchia (W. cf. schneideri, W. piniformis) is also common, followed by the seed ferns Odontopteris (O. subcrenulata, O. osmundaeformis, O. cf. readii), and Autunia (A. conferta, A. naumannii).[9] The plants tend to be adapted to low humidity; macrofossils of moisture-loving plants such as ferns and sphenophytes (horsetails) are very rare, despite the evidence for deposition in a slow stream channel. The most common palynomorphs are Columinisporites ovalis (sphenophyll spores), Potonieisporites spp. (conifer pollen), Colatisporites decorus (indeterminate pollen), and Vesicaspora (seed fern pollen).[9]
Conversely, the middle bed is almost entirely marattialean ferns. These include Pecopteris cf. jongmansii, Aphlebia erdmannii, Asterotheca sp, and several more unnamed species. Sphenophytes such as Sphenophyllum are also present.[9] Vesicaspora and Potonieisporites continue to make up a significant portion of the palynomorphs, though Knoxisporites cf. ruhlandii (indeterminate spores), Cyclogranisporites spp. (marattialean spores), and Punctatisporites spp. (marattialean spores) are even more dominant. The middle bed represents foliage washed into a nearby stagnant pond.[9]
The upper bed has the greatest portion of sphenophytes, not just sphenophylls (Sphenophyllum cf. thonii) but also calamitaleans (Annularia carinata, Calamites). Otherwise, the floral diversity of the upper bed encompasses the same species as the other Kola Switch beds.[9] The upper bed palynoflora is mostly Columinisporites ovalis, Colatisporites decorus, Vesicaspora, Potonieisporites, and the bisaccate pollen Platysaccus cf. saarensis. The combination of fossils from the lower and middle bed is peculiar considering their strongly different depositional environments.[9]
Sanzenbacher Ranch
The Sanzenbacher flora occupies a narrow layer of greyish claystone coarsening upwards to buff-colored siltstone.[41] The most abundant plants are dry-soil species of seed ferns and conifers. Seed fern species include Autunia conferta, Sphenopteris germanica, Neurodontopteris auriculata, Odontopteris subcrenulata, and Rhachiphyllum schenkii, among others. Walchia is the most common conifer, followed by Cordaites.[41] Riparian plants are less common but far from rare, giving Sanzenbacher a 'mixed' ecosystem character similar to, but drier than, the upper bed of Kola Switch.[9] Marattialean ferns such as Pecopteris and Asterotheca are occasionally prevalent. Sphenophytes include Calamites, Annularia spicata, and to a lesser extent species of Sphenophyllum. The most common palynomorphs are seed fern pollen (Vesicaspora, Wilsonites, Anguisporites), while conifer pollen (Potonieisporites) and spores are rarer.[41] Still, the palynomorph diversity of Sanzenbacher is much greater than at Kola Switch.[9]
See also
References
- Martin Sander, P. (1989). "Early Permian depositional environments and pond bonebeds in central Archer County, Texas". Palaeogeography, Palaeoclimatology, Palaeoecology. 69: 1–21. doi:10.1016/0031-0182(89)90153-3. ISSN 0031-0182.
- Tabor, Neil John; Montañez, Isabel P. (2004-06-24). "Morphology and distribution of fossil soils in the Permo-Pennsylvanian Wichita and Bowie Groups, north-central Texas, USA: implications for western equatorial Pangean palaeoclimate during icehouse-greenhouse transition: Permo-Carboniferous palaeoclimate from palaeosols" (PDF). Sedimentology. 51 (4): 851–884. doi:10.1111/j.1365-3091.2004.00655.x.
- Romer, A.S. (1928). "Vertebrate faunal horizons in the Texas Permo-Carboniferous red beds" (PDF). University of Texas Bulletin. 2801: 67–108.
- Romer, A. S. (1935-11-30). "Early history of Texas redbeds vertebrates". Geological Society of America Bulletin. 46 (11): 1597–1657. doi:10.1130/GSAB-46-1597. ISSN 0016-7606.
- Romer, A. S.; Price, L. W. (1940), Review of the Pelycosauria, Geological Society of America Special Papers, vol. 28, Geological Society of America, pp. 1–534, doi:10.1130/spe28-p1
- Romer, Alfred Sherwood (27 November 1974). "The stratigraphy of the Permian Wichita redbeds of Texas". Breviora. 427: 1–31.
- Hentz, Tucker F.; Brown, L.F. Jr (1987), "Wichita Falls-Lawton sheet", Geologic atlas of Texas, Austin: The University of Texas and Bureau of Economic Geology
- Hentz, Tucker F. (1988). "Lithostratigraphy and Paleoenvironments of Upper Paleozoic Continental Red Beds, North-central Texas: Bowie (new) and Wichita (revised) Groups". The University of Texas at Austin, Bureau of Economic Geology, Report of Investigations. 170.
- DiMichele, William A.; Hotton, Carol L.; Looy, Cindy V.; Hook, Robert W. (2019). "Paleoecological and paleoenvironmental interpretation of three successive macrofloras and palynofloras from the Kola Switch locality, lower Permian (Archer City Formation, Bowie Group) of Clay County, Texas, USA". PalZ. 93 (3): 423–451. doi:10.1007/s12542-019-00485-8. ISSN 0031-0220. PMC 8276796. PMID 34262228.
- Lucas, Spencer G. (2006). "Global Permian tetrapod biostratigraphy and biochronology". Geological Society, London, Special Publications. 265 (1): 65–93. doi:10.1144/GSL.SP.2006.265.01.04. ISSN 0305-8719.
- Tabor, Neil J.; DiMichele, William A.; Montañez, Isabel P.; Chaney, Dan S. (2013). "Late Paleozoic continental warming of a cold tropical basin and floristic change in western Pangea" (PDF). International Journal of Coal Geology. 119: 177–186. doi:10.1016/j.coal.2013.07.009. ISSN 0166-5162.
- Wardlaw, Bruce R. (2005). "Age assignment of the Pennsylvanian-Early Permian succession of North Central Texas" (PDF). Permophiles. 46: 21–22.
- DiMichele, William A.; Tabor, Neil J.; Chaney, Dan S.; Nelson, W. John (2006), "From wetlands to wet spots: Environmental tracking and the fate of Carboniferous elements in Early Permian tropical floras", Wetlands through Time, Geological Society of America, doi:10.1130/2006.2399(11), ISBN 978-0-8137-2399-0
- Laurin, Michel (1991). "The osteology of a Lower Permian eosuchian from Texas and a review of diapsid phylogeny". Zoological Journal of the Linnean Society. 101 (1): 59–95. doi:10.1111/j.1096-3642.1991.tb00886.x.
- Reisz, Robert R.; Laurin, Michel; Marjanović, David (2010). "Apsisaurus witteri from the Lower Permian of Texas: yet another small varanopid synapsid, not a diapsid". Journal of Vertebrate Paleontology. 30 (5): 1628–1631. doi:10.1080/02724634.2010.501441. ISSN 0272-4634.
- Romer, A.S. (1937). "New genera and species of pelycosaurian reptiles" (PDF). Proceedings of the New England Zoological Club. 16: 89–97.
- Hook, Robert W. (1989). "Stratigraphic distribution of tetrapods in the Bowie and Wichita Groups, Permo-Carboniferous of north-central Texas". Permo-Carboniferous Vertebrate Paleontology, Lithostratigraphy, and Depositional Environments of North-Central Texas. Field Trip Guidebook No. 2, 49th Annual Meeting of the Society of Vertebrate Paleontology. Austin. pp. 47–53.
{{cite book}}: CS1 maint: location missing publisher (link) - Lucas, Spencer G.; Rinehart, Larry F.; Celeskey, Matthew D. (2018-11-14). "New eupelycosaur". Palaeontologia Electronica. 21 (3): 1–42. doi:10.26879/899. ISSN 1094-8074.
- Llewellyn Ivor Price (1937). "Two new cotylosaurs from the Permian of Texas". Proceedings of the New England Zoölogical Club. 11: 97–102.
- J. Clark; R. L. Carroll (1973). "Romeriid Reptiles from the Lower Permian". Bulletin of the Museum of Comparative Zoology. 144 (5): 353–407.
- Olson, Everett Claire (1941). "The Family Trematopsidae". The Journal of Geology. 49 (2): 149–176. doi:10.1086/624952. ISSN 0022-1376. JSTOR 30080836.
- Carroll, Robert L. (1964). "Early evolution of the dissorophid amphibians". Bulletin of the Museum of Comparative Zoology. 131 (7): 161–250.
- May, William; Huttenlocker, Adam K.; Pardo, Jason D.; Benca, Jeff; Small, Bryan J. (2011). "New Upper Pennsylvanian armored dissorophid records (Temnospondyli, Dissorophoidea) from the U.S. midcontinent and the stratigraphic distributions of dissorophids". Journal of Vertebrate Paleontology. 31 (4): 907–912. doi:10.1080/02724634.2011.582532. ISSN 0272-4634.
- Dilkes, David W. (2020-07-03). "Revision of the Early Permian dissorophid 'Dissorophus' angustus (Temnospondyli: Dissorophoidea)". Journal of Vertebrate Paleontology. 40 (4): e1801704. doi:10.1080/02724634.2020.1801704. ISSN 0272-4634.
- Romer, Alfred S.; Witter, Robert V. (1942). "Edops, a Primitive Rhachitomous Amphibian from the Texas Red Beds". The Journal of Geology. 50 (8): 925–960. doi:10.1086/625101. ISSN 0022-1376.
- Pawley, Kat; Warren, Anne (2006). "The Appendicular Skeleton of Eryops Megacephalus Cope, 1877 (Temnospondyli: Eryopoidea) from the Lower Permian of North America". Journal of Paleontology. 80 (3): 561–580. doi:10.1666/0022-3360(2006)80[561:TASOEM]2.0.CO;2. ISSN 0022-3360.
- Chase, J.N. (1965). "Neldasaurus wrightae, a new rhachitomous labyrinthodont from the Texas Lower Permian". Bulletin of the Museum of Comparative Zoology. 133 (3): 153–225.
- Schoch, Rainer R. (2018-01-01). "Osteology of the temnospondyl Neldasaurus and the evolution of basal dvinosaurians". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 287 (1): 1–16. doi:10.1127/njgpa/2018/0700. ISSN 0077-7749.
- Carroll, Robert L. (21 February 1964). "The relationships of the rhachitomous amphibian Parioxys". American Museum Novitates (2167).
- Gee, Bryan M. (2021-11-08). "Returning to the roots: resolution, reproducibility, and robusticity in the phylogenetic inference of Dissorophidae (Amphibia: Temnospondyli)". PeerJ. 9: e12423. doi:10.7717/peerj.12423. ISSN 2167-8359.
- Maddin, Hillary C.; Fröbisch, Nadia B.; Evans, David C.; Milner, Andrew R. (2013). "Reappraisal of the Early Permian amphibamid Tersomius texensis and some referred material". Comptes Rendus Palevol. 12 (7–8): 447–461. doi:10.1016/j.crpv.2013.06.007.
- Dilkes, David W. (1990-06-21). "A new trematopsid amphibian (Temnospondyli: Dissorophoidea) from the Lower Permian of Texas". Journal of Vertebrate Paleontology. 10 (2): 222–243. doi:10.1080/02724634.1990.10011809. ISSN 0272-4634.
- Milner, Andrew R.; Schoch, Rainer R. (2013-10-01). "Trimerorhachis (Amphibia: Temnospondyli) from the Lower Permian of Texas and New Mexico: cranial osteology, taxonomy and biostratigraphy". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 270 (1): 91–128. doi:10.1127/0077-7749/2013/0360. ISSN 0077-7749.
- Romer, Alfred Sherwood (11 January 1957). "The Appendicular Skeleton of the Permian Embolomerous Amphibian Archeria" (PDF). Contributions from the Museum of Paleontology. 13 (5): 105–159.
- Carroll, Robert L. (1968-11-01). "The postcranial skeleton of the Permian microsaur Pantylus". Canadian Journal of Zoology. 46 (6): 1175–1192. doi:10.1139/z68-168. ISSN 0008-4301.
- Carlson, Keith J. (1999). "Crossotelos, an Early Permian nectridian amphibian". Journal of Vertebrate Paleontology. 19 (4): 623–631. doi:10.1080/02724634.1999.10011176. ISSN 0272-4634. JSTOR 4524033.
- Zidek, Jiri (1978). "New Chondrichthyan Spines from the Late Paleozoic of Oklahoma". Journal of Paleontology. 52 (5): 1070–1078. JSTOR 1303852.
- Romer, A. S. (1942-03-01). "Notes on certain American Paleozoic fishes". American Journal of Science. 240 (3): 216–228. doi:10.2475/ajs.240.3.216. ISSN 0002-9599.
- Johnson, Gary D. (2012). "Possible origin of the xenacanth sharks Orthacanthus texensis and Orthacanthus platypternus in the Lower Permian of Texas, USA". Historical Biology. 24 (4): 369–379. doi:10.1080/08912963.2012.669128. ISSN 0891-2963.
- Thomson, Keith Stewart (1964). "Revised generic diagnoses of the fossil fishes Megalichthys and Ectosteorhachis (family Osteolepidae)". Bulletin of the Museum of Comparative Zoology at Harvard University. 131 (9): 283–311.
- DiMichele, William A.; Hook, Robert W.; Kerp, Hans; Hotton, Carol L.; Looy, Cindy V.; Chaney, Dan S. (2018), "Lower Permian Flora of the Sanzenbacher Ranch, Clay County, Texas" (PDF), Transformative Paleobotany, Elsevier, pp. 95–126, doi:10.1016/B978-0-12-813012-4.00006-1, ISBN 9780128130124
- Various Contributors to the Paleobiology Database. "Fossilworks: Gateway to the Paleobiology Database". Retrieved 17 December 2021.