BOK (gene)
Bok (Bcl-2 related ovarian killer) is a protein-coding gene of the Bcl-2 family that is found in many invertebrates and vertebrates. It induces apoptosis, a special type of cell death. Currently, the precise function of Bok in this process is unknown.

Discovery and homology
In 1997, the protein Bcl-2-related ovarian killer (Bok) was identified in a yeast two-hybrid experiment with a rat ovarian cDNA library in a screen for proteins interacting with Mcl-1, an abundant anti-apoptotic protein.[5] The overexpression of Bok induces apoptosis. Because of its high sequence similarity to Bak and Bax,[6] Bok is classified as a member of the Bcl-2 protein family.[7] The mouse homologue of Bok is called Matador (Mtd). This name is derived from the Latin term mactator which means butcher or killer.[8] Additionally, homologous proteins were found in Drosophila melanogaster (fruit fly) and Gallus gallus (chicken).[9]
Promoter and gene structure
The human BOK promoter is activated by the overexpression of members of the E2F hand transcription factor family. Typically, these transcription factors are involved in the promotion of S-phase, so there might be a connection between Bok expression and cell cycle progression.[10] Due to this regulation of Bok expression by the cell cycle, it was proposed that Bok sensitizes growing cells to stress-induced apoptosis.[10]
Bok mRNA comprises five exons which code for a 213 amino acid protein,[7] called Bok-L. This protein consists of four Bcl-2 homology domains (abbreviated BH1, BH2, BH3, BH4, respectively) and a C-terminal transmembrane region [7] (Figure 1). Its BH3 domain contains a stretch with many leucine residues. This is unique among the Bcl-2 family members. The leucine-rich stretch functions as a nuclear export signal.[11] It is recognized by the nuclear exportin Crm1. Mutations in the leucine-rich stretch impair the binding of Crm1 to Bok.[11] Consequently, Bok accumulates in the nucleus and triggers apoptosis.[11]
Splice variants
Due to alternative splicing, Bok mRNA gives rise to different Bok proteins: Figure 1 illustrates the different splice variants schematically. Full length Bok is named Bok-L.
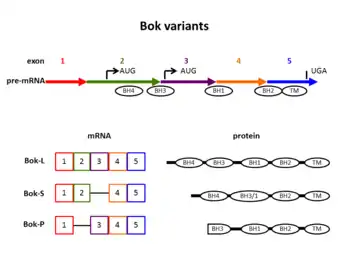
The shorter version, Bok-S, lacks exon 3. This results in a fusion of the BH3 domain with the BH1 domain.[6][12] The BH3 domain is involved in the interaction of Bok with Mcl-1 and other molecules. It is dispensable for the induction of apoptosis.[12] Expression of Bok-S may be an immediate response to stress signals. It has been shown to induce apoptosis regardless of the presence of anti-apoptotic molecules.[6]
Another splice variant termed Bok-P was found in placental tissue from patients with pre-eclampsia. While Bok-S misses exon 3, Bok-P lacks exon 2. This deletion includes the BH4 domain and parts of the BH3 domain. Bok-P may be the cause for trophoblast cell death in pre-eclampsia,[7] a dangerous pregnancy complication. In pre-eclampsia, typical alterations occur in the maternal kidney and lead to hypertension and proteins in the urine. To date, the cause of this medical condition as well as an appropriate treatment have not been discovered.
Expression pattern
The Bok gene is activated and produces protein in different tissues. In mice, elevated Bok levels were detected in the ovary, the testis, and the uterus.[5] Nevertheless, it also exists in the brain and at low levels in most other tissues.[13] However, the expression pattern of the Bok gene varies among species.
In humans, Bok is found in a wide range of tissues. The gene is expressed in the colon, the stomach, the testes, the placenta, the pancreas, the ovaries, and the uterus.[14] Furthermore, more Bok is expressed in fetal tissue compared to adult tissue. Thus, Bok may influence development.
Subcellular localization
The subcellular localization of Bok protein is controversial. In proliferating cells, Bok is found in the nucleus.[15] Upon induction of apoptosis, it was found to tightly associate with mitochondrial membranes.[14][15] On the other hand, another group found Bok shuttling between the cytoplasm and the nucleus. In their experiments, increased nuclear (not mitochondrial) localization correlated with a stronger apoptotic activity.[11]
Regulation
It was found that the cellular ratio of pro-apoptotic to anti-apoptotic Bcl-2 family members effects late apoptotic events such as release of cytochrome c from the mitochondria and the activation of caspases. Higher levels of pro-apoptotic proteins compared to anti-apoptotic proteins seem to cause apoptosis. In a current model, the formation of heterodimers between pro-apoptotic and anti-apoptotic proteins prevents induction of apoptosis.[12]
Interactions
The binding of Bok to its interacting partners seems to be mediated by its BH3 domain.[7] The splice variant Bok-S lacks this domain and is unable to form heterodimers with other proteins of the Bcl-2 family.
In yeast two-hybrid experiments, Bok was found to interact with the anti-apoptotic proteins Mcl-1, BHRF-1, and Bfl-1. However, interactions with other anti-apoptotic proteins such as Bcl-2, Bcl-xL, and Bcl-w were not detectable (1). Later studies aimed at confirming an interaction between Bok and pro-apoptotic Bak or Bax but were not successful.[8]
Accordingly, coexpression of anti-apoptotic proteins such as Mcl-1 suppresses apoptosis induced by Bok overexpression.[5] Consistent with the results mentioned above, coexpression of anti-apoptotic Bcl-2 does not prevent Bok-induced apoptosis.[5]
Knock-out mouse
Since its discovery in 1997, several attempts have been made to characterize Bok. Due to the increased expression levels in fetal tissue, scientists anticipated a developmental role for Bok. Recently, the Bok knock-out mouse was created. This mouse shows, however, no developmental defects and normal fertility.[13] This finding indicates that the function of Bok seems to overlap with the function of the related pro-apoptotic proteins Bak and Bax.
Several other roles were proposed for Bok, especially in developing cells.[12][16][17][18] Since the action of Bok in triggering apoptosis seems to be redundant, it is difficult to assign a specific role to Bok in the presence of Bak and Bax. The study of cells deficient in Bak and Bok or deficient in Bax and Bok, respectively, could help to better characterize the role of Bok in apoptosis. If Bok exerts a critical function, it is likely that this function is limited to certain circumstances, e.g. specific cell types, stress conditions. Thus, these aspects should be assessed in more detail to analyze the physiological and pathological role of Bok.
References
- GRCh38: Ensembl release 89: ENSG00000176720 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000026278 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Hsu SY, Kaipia A, McGee E, Lomeli M, Hsueh AJ (November 1997). "Bok is a pro-apoptotic Bcl-2 protein with restricted expression in reproductive tissues and heterodimerizes with selective anti-apoptotic Bcl-2 family members". Proceedings of the National Academy of Sciences of the United States of America. 94 (23): 12401–6. Bibcode:1997PNAS...9412401H. doi:10.1073/pnas.94.23.12401. PMC 24966. PMID 9356461.
- Hsu SY, Hsueh AJ (November 1998). "A splicing variant of the Bcl-2 member Bok with a truncated BH3 domain induces apoptosis but does not dimerize with antiapoptotic Bcl-2 proteins in vitro". The Journal of Biological Chemistry. 273 (46): 30139–46. doi:10.1074/jbc.273.46.30139. PMID 9804769.
- Soleymanlou N, Wu Y, Wang JX, Todros T, Ietta F, Jurisicova A, Post M, Caniggia I (May 2005). "A novel Mtd splice isoform is responsible for trophoblast cell death in pre-eclampsia". Cell Death and Differentiation. 12 (5): 441–52. doi:10.1038/sj.cdd.4401593. PMID 15775999.
- Inohara N, Ekhterae D, Garcia I, Carrio R, Merino J, Merry A, Chen S, Núñez G (April 1998). "Mtd, a novel Bcl-2 family member activates apoptosis in the absence of heterodimerization with Bcl-2 and Bcl-XL". The Journal of Biological Chemistry. 273 (15): 8705–10. doi:10.1074/jbc.273.15.8705. PMID 9535847.
- Zhang H, Holzgreve W, De Geyter C (September 2000). "Evolutionarily conserved Bok proteins in the Bcl-2 family". FEBS Letters. 480 (2–3): 311–3. doi:10.1016/s0014-5793(00)01921-9. PMID 11034351. S2CID 19422343.
- Rodriguez JM, Glozak MA, Ma Y, Cress WD (August 2006). "Bok, Bcl-2-related Ovarian Killer, Is Cell Cycle-regulated and Sensitizes to Stress-induced Apoptosis". The Journal of Biological Chemistry. 281 (32): 22729–35. doi:10.1074/jbc.M604705200. PMC 2134790. PMID 16772296.
- Bartholomeusz G, Wu Y, Ali Seyed M, Xia W, Kwong KY, Hortobagyi G, Hung MC (February 2006). "Nuclear translocation of the pro-apoptotic Bcl-2 family member Bok induces apoptosis". Molecular Carcinogenesis. 45 (2): 73–83. doi:10.1002/mc.20156. PMID 16302269. S2CID 24230787.
- Hsu SY, Hsueh AJ (April 2000). "Tissue-specific Bcl-2 protein partners in apoptosis: An ovarian paradigm". Physiological Reviews. 80 (2): 593–614. doi:10.1152/physrev.2000.80.2.593. PMID 10747202. S2CID 6203924.
- Ke F, Voss A, Kerr JB, O'Reilly LA, Tai L, Echeverry N, Bouillet P, Strasser A, Kaufmann T (June 2012). "BCL-2 family member BOK is widely expressed but its loss has only minimal impact in mice". Cell Death and Differentiation. 19 (6): 915–25. doi:10.1038/cdd.2011.210. PMC 3354060. PMID 22281706.
- Gao S, Fu W, Dürrenberger M, De Geyter C, Zhang H (May 2005). "Membrane translocation and oligomerization of hBok are triggered in response to apoptotic stimuli and Bnip3" (PDF). Cellular and Molecular Life Sciences. 62 (9): 1015–24. doi:10.1007/s00018-005-4543-3. PMID 15868100. S2CID 2947.
- Ray JE, Garcia J, Jurisicova A, Caniggia I (May 2010). "Mtd/Bok takes a swing: proapoptotic Mtd/Bok regulates trophoblast cell proliferation during human placental development and in preeclampsia". Cell Death and Differentiation. 17 (5): 846–59. doi:10.1038/cdd.2009.167. PMID 19942931.
- Suominen JS, Yan W, Toppari J, Kaipia A (December 2001). "The expression and regulation of Bcl-2-related ovarian killer (Bok) mRNA in the developing and adult rat testis". European Journal of Endocrinology. 145 (6): 771–8. doi:10.1530/eje.0.1450771. PMID 11720903.
- Soane L, Siegel ZT, Schuh RA, Fiskum G (May 2008). "Postnatal developmental regulation of Bcl-2 family proteins in brain mitochondria". Journal of Neuroscience Research. 86 (6): 1267–76. doi:10.1002/jnr.21584. PMC 2566804. PMID 18058945.
- Jääskeläinen M, Nieminen A, Pökkylä RM, Kauppinen M, Liakka A, Heikinheimo M, Vaskivuo TE, Klefström J, Tapanainen JS (December 2010). "Regulation of cell death in human fetal and adult ovaries--role of Bok and Bcl-X(L)" (PDF). Molecular and Cellular Endocrinology. 330 (1–2): 17–24. doi:10.1016/j.mce.2010.07.020. PMID 20673843. S2CID 1972679.
External links
- Human BOK genome location and BOK gene details page in the UCSC Genome Browser.