Arrestin
Arrestins (abbreviated Arr) are a small family of proteins important for regulating signal transduction at G protein-coupled receptors.[2][3] Arrestins were first discovered as a part of a conserved two-step mechanism for regulating the activity of G protein-coupled receptors (GPCRs) in the visual rhodopsin system by Hermann Kühn, Scott Hall, and Ursula Wilden[4] and in the β-adrenergic system by Martin J. Lohse and co-workers.[5][6]
| S-antigen; retina and pineal gland (arrestin) | |||||||
|---|---|---|---|---|---|---|---|
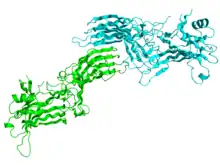 Crystallographic structure of the bovine arrestin-S.[1] | |||||||
| Identifiers | |||||||
| Symbol | SAG | ||||||
| Alt. symbols | arrestin-1 | ||||||
| NCBI gene | 6295 | ||||||
| HGNC | 10521 | ||||||
| OMIM | 181031 | ||||||
| RefSeq | NM_000541 | ||||||
| UniProt | P10523 | ||||||
| Other data | |||||||
| Locus | Chr. 2 q37.1 | ||||||
| |||||||
| arrestin beta 1 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | ARRB1 | ||||||
| Alt. symbols | ARR1, arrestin-2 | ||||||
| NCBI gene | 408 | ||||||
| HGNC | 711 | ||||||
| OMIM | 107940 | ||||||
| RefSeq | NM_004041 | ||||||
| UniProt | P49407 | ||||||
| Other data | |||||||
| Locus | Chr. 11 q13 | ||||||
| |||||||
| arrestin beta 2 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | ARRB2 | ||||||
| Alt. symbols | ARR2, arrestin-3 | ||||||
| NCBI gene | 409 | ||||||
| HGNC | 712 | ||||||
| OMIM | 107941 | ||||||
| RefSeq | NM_004313 | ||||||
| UniProt | P32121 | ||||||
| Other data | |||||||
| Locus | Chr. 17 p13 | ||||||
| |||||||
| arrestin 3, retinal (X-arrestin) | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | ARR3 | ||||||
| Alt. symbols | ARRX, arrestin-4 | ||||||
| NCBI gene | 407 | ||||||
| HGNC | 710 | ||||||
| OMIM | 301770 | ||||||
| RefSeq | NM_004312 | ||||||
| UniProt | P36575 | ||||||
| Other data | |||||||
| Locus | Chr. X q | ||||||
| |||||||
Function
In response to a stimulus, GPCRs activate heterotrimeric G proteins. In order to turn off this response, or adapt to a persistent stimulus, active receptors need to be desensitized. The first step in desensitization is phosphorylation of the receptor by a class of serine/threonine kinases called G protein coupled receptor kinases (GRKs). GRK phosphorylation specifically prepares the activated receptor for arrestin binding. Arrestin binding to the receptor blocks further G protein-mediated signaling and targets receptors for internalization, and redirects signaling to alternative G protein-independent pathways, such as β-arrestin signaling.[7][8][9][10][6] In addition to GPCRs, arrestins bind to other classes of cell surface receptors and a variety of other signaling proteins.[11]
Subtypes
Mammals express four arrestin subtypes and each arrestin subtype is known by multiple aliases. The systematic arrestin name (1-4) plus the most widely used aliases for each arrestin subtype are listed in bold below:
- Arrestin-1 was originally identified as the S-antigen (SAG) causing uveitis (autoimmune eye disease), then independently described as a 48 kDa protein that binds light-activated phosphorylated rhodopsin before it became clear that both are one and the same. It was later renamed visual arrestin, but when another cone-specific visual subtype was cloned the term rod arrestin was coined. This also turned out to be a misnomer: arrestin-1 expresses at comparable very high levels in both rod and cone photoreceptor cells.
- Arrestin-2 was the first non-visual arrestin cloned. It was first named β-arrestin simply because of the two GPCRs available in purified form at the time, rhodopsin and β2-adrenergic receptor, it showed preference for the latter.
- Arrestin-3. The second non-visual arrestin cloned was first termed β-arrestin-2 (retroactively changing the name of β-arrestin into β-arrestin-1), even though by that time it was clear that non-visual arrestins interact with hundreds of different GPCRs, not just with β2-adrenergic receptor. Systematic names, arrestin-2 and arrestin-3, respectively, were proposed soon after that.
- Arrestin-4 was cloned by two groups and termed cone arrestin, after photoreceptor type that expresses it, and X-arrestin, after the chromosome where its gene resides. In the HUGO database its gene is called arrestin-3.
Fish and other vertebrates appear to have only three arrestins: no equivalent of arrestin-2, which is the most abundant non-visual subtype in mammals, was cloned so far. The proto-chordate Ciona intestinalis (sea squirt) has only one arrestin, which serves as visual in its mobile larva with highly developed eyes, and becomes generic non-visual in the blind sessile adult. Conserved positions of multiple introns in its gene and those of our arrestin subtypes suggest that they all evolved from this ancestral arrestin.[12] Lower invertebrates, such as roundworm Caenorhabditis elegans, also have only one arrestin. Insects have arr1 and arr2, originally termed “visual arrestins” because they are expressed in photoreceptors, and one non-visual subtype (kurtz in Drosophila). Later arr1 and arr2 were found to play an important role in olfactory neurons and renamed “sensory”. Fungi have distant arrestin relatives involved in pH sensing.
Tissue distribution
One or more arrestin is expressed in virtually every eukaryotic cell. In mammals, arrestin-1 and arrestin-4 are largely confined to photoreceptors, whereas arrestin-2 and arrestin-3 are ubiquitous. Neurons have the highest expression level of both non-visual subtypes. In neuronal precursors both are expressed at comparable levels, whereas in mature neurons arrestin-2 is present at 10-20 fold higher levels than arrestin-3.
Mechanism
Arrestins block GPCR coupling to G proteins in two ways. First, arrestin binding to the cytoplasmic face of the receptor occludes the binding site for heterotrimeric G-protein, preventing its activation (desensitization).[13] Second, arrestin links the receptor to elements of the internalization machinery, clathrin and clathrin adaptor AP2, which promotes receptor internalization via coated pits and subsequent transport to internal compartments, called endosomes. Subsequently, the receptor could be either directed to degradation compartments (lysosomes) or recycled back to the plasma membrane where it can again signal. The strength of arrestin-receptor interaction plays a role in this choice: tighter complexes tend to increase the probability of receptor degradation (Class B), whereas more transient complexes favor recycling (Class A), although this “rule” is far from absolute.[2] More recently direct interactions between Gi/o family G proteins and Arrestin were discovered downstream of multiple receptors, regardless of canonical G protein coupling.[14] These recent findings introduce a GPCR signaling mechanism distinct from canonical G protein activation and β-arrestin desensitization in which GPCRs cause the formation of Gαi:β-arrestin signaling complexes.
Structure
Arrestins are elongated molecules, in which several intra-molecular interactions hold the relative orientation of the two domains. Unstimulated cell arrestins are localized in the cytoplasm in a basal “inactive” conformation. Active phosphorylated GPCRs recruit arrestin to the plasma membrane. Receptor binding induces a global conformational change that involves the movement of the two arrestin domains and the release of its C-terminal tail that contains clathrin and AP2 binding sites. Increased accessibility of these sites in receptor-bound arrestin targets the arrestin-receptor complex to the coated pit. Arrestins also bind microtubules (part of the cellular “skeleton”), where they assume yet another conformation, different from both free and receptor-bound form. Microtubule-bound arrestins recruit certain proteins to the cytoskeleton, which affects their activity and/or redirects it to microtubule-associated proteins.
Arrestins shuttle between cell nucleus and cytoplasm. Their nuclear functions are not fully understood, but it was shown that all four mammalian arrestin subtypes remove some of their partners, such as protein kinase JNK3 or the ubiquitin ligase Mdm2, from the nucleus. Arrestins also modify gene expression by enhancing transcription of certain genes.
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

References
- PDB: 1CF1; Hirsch JA, Schubert C, Gurevich VV, Sigler PB (April 1999). "The 2.8 A crystal structure of visual arrestin: a model for arrestin's regulation". Cell. 97 (2): 257–69. doi:10.1016/S0092-8674(00)80735-7. PMID 10219246. S2CID 17124300.
- Moore CA, Milano SK, Benovic JL (2007). "Regulation of receptor trafficking by GRKs and arrestins". Annual Review of Physiology. 69: 451–82. doi:10.1146/annurev.physiol.69.022405.154712. PMID 17037978.
- Lefkowitz RJ, Shenoy SK (April 2005). "Transduction of receptor signals by beta-arrestins". Science. 308 (5721): 512–7. Bibcode:2005Sci...308..512L. doi:10.1126/science.1109237. PMID 15845844. S2CID 26931077.
- Wilden U, Hall SW, Kühn H (March 1986). "Phosphodiesterase activation by photoexcited rhodopsin is quenched when rhodopsin is phosphorylated and binds the intrinsic 48-kDa protein of rod outer segments". Proceedings of the National Academy of Sciences of the United States of America. 83 (5): 1174–8. Bibcode:1986PNAS...83.1174W. doi:10.1073/pnas.83.5.1174. PMC 323037. PMID 3006038.
- Lohse MJ, Benovic JL, Codina J, Caron MG, Lefkowitz RJ (June 1990). "beta-Arrestin: a protein that regulates beta-adrenergic receptor function". Science. 248 (4962): 1547–50. Bibcode:1990Sci...248.1547L. doi:10.1126/science.2163110. PMID 2163110.
- Gurevich VV, Gurevich EV (June 2006). "The structural basis of arrestin-mediated regulation of G-protein-coupled receptors". Pharmacology & Therapeutics. 110 (3): 465–502. doi:10.1016/j.pharmthera.2005.09.008. PMC 2562282. PMID 16460808.
- Smith JS, Lefkowitz RJ, Rajagopal S (January 2018). "Biased signalling: from simple switches to allosteric microprocessors". Nature Reviews. Drug Discovery. 17 (4): 243–260. doi:10.1038/nrd.2017.229. PMC 5936084. PMID 29302067.
- Cahill TJ, Thomsen AR, Tarrasch JT, Plouffe B, Nguyen AH, Yang F, et al. (February 2017). "Distinct conformations of GPCR-β-arrestin complexes mediate desensitization, signaling, and endocytosis". Proceedings of the National Academy of Sciences of the United States of America. 114 (10): 2562–2567. Bibcode:2017PNAS..114.2562C. doi:10.1073/pnas.1701529114. PMC 5347553. PMID 28223524.
- Kumari P, Srivastava A, Banerjee R, Ghosh E, Gupta P, Ranjan R, Chen X, Gupta B, Gupta C, Jaiman D, Shukla AK (November 2016). "Functional competence of a partially engaged GPCR-β-arrestin complex". Nature Communications. 7: 13416. Bibcode:2016NatCo...713416K. doi:10.1038/ncomms13416. PMC 5105198. PMID 27827372.
- Kumari P, Srivastava A, Ghosh E, Ranjan R, Dogra S, Yadav PN, Shukla AK (April 2017). "Core engagement with β-arrestin is dispensable for agonist-induced vasopressin receptor endocytosis and ERK activation". Molecular Biology of the Cell. 28 (8): 1003–10. doi:10.1091/mbc.E16-12-0818. PMC 5391177. PMID 28228552.
- Gurevich VV, Gurevich EV (February 2004). "The molecular acrobatics of arrestin activation". Trends in Pharmacological Sciences. 25 (2): 105–11. doi:10.1016/j.tips.2003.12.008. PMID 15102497.
- Gurevich EV, Gurevich VV (2006). "Arrestins: ubiquitous regulators of cellular signaling pathways". Genome Biology. 7 (9): 236. doi:10.1186/gb-2006-7-9-236. PMC 1794542. PMID 17020596.
- Kang Y, Zhou XE, Gao X, He Y, Liu W, Ishchenko A, et al. (July 2015). "Crystal structure of rhodopsin bound to arrestin by femtosecond X-ray laser". Nature. 523 (7562): 561–7. Bibcode:2015Natur.523..561K. doi:10.1038/nature14656. PMC 4521999. PMID 26200343.
- Smith JS, Pack TF, et al. (2021). "Noncanonical scaffolding of Gαi and β-arrestin by G protein–coupled receptors". Science. 371 (Ahead of print): eaay1833. doi:10.1126/science.aay1833. PMC 8005335. PMID 33479120.
- Han M, Gurevich VV, Vishnivetskiy SA, Sigler PB, Schubert C (September 2001). "Crystal structure of beta-arrestin at 1.9 A: possible mechanism of receptor binding and membrane Translocation". Structure. 9 (9): 869–80. doi:10.1016/S0969-2126(01)00644-X. PMID 11566136.
External links
- Arrestin at the U.S. National Library of Medicine Medical Subject Headings (MeSH)