Calcium encoding
Calcium encoding (also referred to as Ca2+ encoding or calcium information processing) is an intracellular signaling pathway used by many cells to transfer, process and encode external information detected by the cell. In cell physiology, external information is often converted into intracellular calcium dynamics. The concept of calcium encoding explains how Ca2+ ions act as intracellular messengers, relaying information within cells to regulate their activity.[1] Given the ubiquity of Ca2+ ions in cell physiology, Ca2+ encoding has also been suggested as a potential tool to characterize cell physiology in health and disease.[2][3][4] The mathematical bases of Ca2+ encoding have been pioneered by work of Joel Keizer and Hans G. Othmer on calcium modeling in the 1990s and more recently they have been revisited by Eshel Ben-Jacob, Herbert Levine and co-workers.
AM, FM and AFM calcium encoding
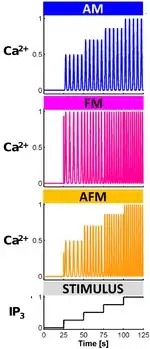
Although elevations of Ca2+ are necessary for it to act as a signal, prolonged increases of the concentration of Ca2+ in the cytoplasm can be lethal for the cell. Thus cells avoid death usually delivering Ca2+ signals as brief transients - i.e. Ca2+ elevations followed by a rapid decay - or in the form of oscillations. In analogy with Information Theory, either the amplitude or the frequency or both features of these Ca2+ oscillations define the Ca2+ encoding mode. Therefore, three classes of Ca2+ signals can be distinguished on the basis of their encoding mode:[2][3][5]
- AM encoding Ca2+ signals: when the strength of the stimuli is encoded by amplitude modulations of Ca2+ oscillations. In other words, stimuli of the same nature but of different strength are associated with different amplitudes of Ca2+ oscillations but the frequencies of these oscillations are similar;
- FM encoding Ca2+ signals: when the strength of the stimuli is encoded by frequency modulations of Ca2+ oscillations. In other words, stimuli of the same nature but of different strength are associated with different frequencies of Ca2+ oscillations but the amplitudes of these oscillations are similar;
- AFM encoding Ca2+ signals: when both AM and FM encoding modes coexist.
Experiments and biophysical modeling, show that the mode of calcium encoding varies from cell to cell, and that a given cell could even show different types of calcium encoding for different patho-physiological conditions.[6][7] This could ultimately provide a crucial tool in medical diagnostics, to characterize, recognize and prevent diseases.[6] Analysis tools based on Furier transform have been developed that can perform spectral analysis and examine calcium oscillations from experimental data [8]
Mathematical aspects of calcium encoding
Calcium encoding can be mathematically characterized by biophysical models of calcium signaling.[9] Phase plane and bifurcation analysis of these models, can reveal in fact how frequency and amplitude of calcium oscillations vary as a function of any parameter of the model.[3] Occurrence of AM-, FM- or AFM-encoding can be assessed on the extension of the min-max range of amplitude and frequency of Ca2+ oscillations and the bifurcation structure of the system under study.
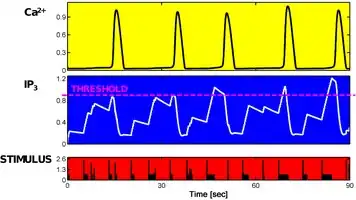
A critical aspect of Ca2+ encoding revealed by modeling, is how it depends on the dynamics of the complex network of reactions of underlying Ca2+-mobilizing signals. This aspect can be addressed considering Ca2+ models that include both Ca2+ dynamics and the dynamics of Ca2+-mobilizing signals. One simple and biophysically realistic model of this kind is the ChI model, originally developed by Eshel Ben-Jacob and co-workers,[4] for GPCR-mediated inositol-1,4,5-trisphosphate (IP3)-triggered Ca2+-induced Ca2+-release. The main conclusion of this study was that the dynamics of the Ca2+-mobilizing IP3 signal is essentially AFM encoding with respect to the stimulus whereas Ca2+ oscillations can be either FM or AFM but not solely AM.[4] It was argued that the AFM nature of the Ca2+-mobilizing IP3 signal could represent the ideal solution to optimally translate pulsed or discontinuous extracellular signals into intracellular continuous-like Ca2+ oscillations.[4]
Computational aspects
Calcium encoding can be confined within a single cell or involve cell ensembles[9][10] and deploy essential computational tasks, such as stimulus integration[4][11] or regulated activation of gene transcription.[12] Moreover, cells are often organized in networks, allowing intercellular propagation of calcium signaling.[10] With this regard, the same mode of calcium encoding could be shared by different cells, providing synchronization[4] or the functional basis to implement more complex computational tasks.[7]
Michael Forrest has shown that intracellular calcium dynamics may permit a Purkinje neuron to perform toggle and gain computations upon its inputs.[13] So, showing how an ion concentration can be used as a computational variable - in particular, as a memory element: recording a history of firing and inputs, to dictate how the neuron responds to future inputs. Thus, this work hypothesizes that the membrane potential (V) is not the Purkinje cell's only coding variable but works alongside a calcium memory system. These two interact, with the calcium memory being encoded and decoded by the membrane potential. The toggle and gain computations are likely to be salient to network computations in the cerebellum, in the brain. Thus, ion computations can be important more globally than a single cell. Forrest terms this hypothesis: "ion to network computation".
References
- Berridge, M. J.; Bootman, M. D.; Lipp, P. (1998). "Calcium – a life and death signal". Nature. 395 (6703): 645–648. Bibcode:1998Natur.395..645B. doi:10.1038/27094. PMID 9790183.
- De Pittà, M.; Volman, V.; Levine, H.; Ben-Jacob, E. (2009). "Multimodal encoding in a simplified model of intracellular calcium signaling". Cognitive Processing. 10 (S1): 55–70. arXiv:0912.3064. Bibcode:2009arXiv0912.3064D. doi:10.1007/s10339-008-0242-y. PMID 19030904. S2CID 91230.
- De Pittà, M.; Volman, V.; Levine, H.; Pioggia, G.; De Rossi, D.; Ben-Jacob, E. (2008). "Coexistence of amplitude and frequency modulations in intracellular calcium dynamics". Phys. Rev. E. 77 (3): 030903(R). arXiv:0806.2373. Bibcode:2008PhRvE..77c0903D. doi:10.1103/PhysRevE.77.030903. PMID 18517322. S2CID 17221301.
- De Pittà, M.; Goldberg, M.; Volman, V.; Berry, H.; Ben-Jacob, E. (2009). "Glutamate-dependent intracellular calcium and IP3 oscillating and pulsating dynamics in astrocytes". J. Biol. Phys. 35 (4): 383–411. arXiv:0912.3057. doi:10.1007/s10867-009-9155-y. PMC 2750743. PMID 19669422.
- Berridge, M.J. (1997). "The AM and FM of calcium signaling". Nature. 386 (6627): 759–760. Bibcode:1997Natur.386..759B. doi:10.1038/386759a0. PMID 9126727.
- Berridge, M. J.; Bootman, M. D.; Roderick, H. L. (2003). "Calcium signalling: dynamics, homeostasis and remodelling" (PDF). Nature Reviews Molecular Cell Biology. 4 (7): 517–529. doi:10.1038/nrm1155. PMID 12838335. S2CID 1152297.
- Goldberg, M.; De Pittà, M.; Volman, V.; Berry, H.; Ben-Jacob, E. (2010). "Nonlinear gap junctions enable long-distance propagation of pulsating calcium waves in astrocyte networks". PLOS Comput. Biol. 6 (8): e1000909. arXiv:1009.2243. Bibcode:2010PLSCB...6E0909G. doi:10.1371/journal.pcbi.1000909. PMC 2928752. PMID 20865153.
- Uhlen, P. (2004). "Spectral Analysis of Calcium Oscillations". Science's STKE. 2004 (258): pl15. doi:10.1126/stke.2582004pl15. PMID 15536176.
- Falcke, M. (2004). "Reading the patterns in living cells – The physics of Ca2+ signaling". Adv. Phys. 53 (3): 255–440. Bibcode:2004AdPhy..53..255F. doi:10.1080/00018730410001703159. S2CID 119764503.
- Scemes, E.; Giaume, C. (2006). "Astrocyte calcium waves: What they are and what they do". Glia. 54 (7): 716–725. doi:10.1002/glia.20374. PMC 2605018. PMID 17006900.
- Tang, Y.; Othmer, H. G. (1995). "Frequency encoding in excitable systems with applications to calcium oscillations". Proc. Natl. Acad. Sci. USA. 92 (17): 7869–7873. Bibcode:1995PNAS...92.7869T. doi:10.1073/pnas.92.17.7869. PMC 41247. PMID 7644505.
- Dolmetsch, R. E.; Lewis, R. S.; Goodnow, C. C.; Healy, J. I. (1997). "Differential activation of transcription factors induced by Ca2+ response amplitude and duration". Nature. 386 (24): 855–858. Bibcode:1997Natur.386..855D. doi:10.1038/386855a0. PMID 9126747. S2CID 4366554.
- Forrest MD (2014). "Intracellular Calcium Dynamics Permit a Purkinje Neuron Model to Perform Toggle and Gain Computations Upon its Inputs". Frontiers in Computational Neuroscience. 8: 86. doi:10.3389/fncom.2014.00086. PMC 4138505. PMID 25191262.