Carboxysome
Carboxysomes are bacterial microcompartments (BMCs) consisting of polyhedral protein shells filled with the enzymes ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO)—the predominant enzyme in carbon fixation and the rate limiting enzyme in the Calvin cycle—and carbonic anhydrase.[2]
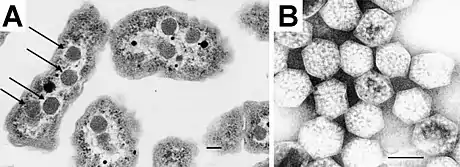
Carboxysomes are thought to have evolved as a consequence of the increase in oxygen concentration in the ancient atmosphere; this is because oxygen is a competing substrate to carbon dioxide in the RuBisCO reaction.[3] To overcome the inefficiency of RuBisCO, carboxysomes concentrate carbon dioxide inside the shell by means of co-localized carbonic anhydrase activity, which produces carbon dioxide from the bicarbonate that diffuses into the carboxysome. The resulting concentration of carbon dioxide near RuBisCO decreases the proportion of ribulose-1,5-bisphosphate oxygenation and thereby avoids costly photorespiratory reactions. The surrounding shell provides a barrier to carbon dioxide loss, helping to increase its concentration around RuBisCO.[4][5][6]
Carboxysomes are an essential part of the broader metabolic network called the Carbon dioxide-Concentrating Mechanism (CCM), which functions in two parts:[7] (1) Membrane transporters concentrate inorganic carbon (Ci) in the cell cytosol which is devoid of carbonic anhydrases. Carbon is primarily stored in the form of HCO3- which cannot re-cross the lipid membrane, as opposed to neutral CO2 which can easily escape the cell. This stockpiles carbon in the cell, creating a disequilibrium between the intracellular and extracellular environments of about 30x the Ci concentration in water.[8] (2) Cytosolic HCO3- diffuses into the carboxysome, where carboxysomal carbonic anhydrases dehydrate it back to CO2 in the vicinity of Rubisco, allowing Rubisco to operate at its maximal rate.
Carboxysomes are the best studied example of bacterial microcompartments, the term for functionally diverse organelles that are alike in having a protein shell.[9][10]
Discovery
Polyhedral bodies were discovered by transmission electron microscopy in the cyanobacterium Phormidium uncinatum in 1956.[11] These were later observed in other cyanobacteria[12] and in some chemotrophic bacteria that fix carbon dioxide—many of them are sulfur oxidizers or nitrogen fixers (for example, Halothiobacillus, Acidithiobacillus, Nitrobacter and Nitrococcus; all belonging to Pseudomonadota).[2][13] The polyhedral bodies were first purified from Thiobacillus neapolitanus (now Halothiobacillus neapolitanus) in 1973 and shown to contain RuBisCO, held within a rigid outer covering.[14] The authors proposed that since these appeared to be organelles involved in carbon dioxide fixation, they should be called carboxysomes.[14]
Architecture

Structurally, carboxysomes are icosahedral, or quasi-icosahedral. Electron cryo-tomography studies[15][16][17] have confirmed the approximately icosahedral geometry of the carboxysome, and have imaged Rubisco proteins inside arranged in a few concentric layers or fibril-like structures.[15][17][18] The non-icosahedral faceted shapes of some carboxysomes can naturally be explained within the elastic theory of heterogeneous thin shells.[19]
Shell Proteins
The carboxysome has an outer shell composed of a few thousand protein subunits, with hexameric shell proteins populating the faces and pentameric shell proteins placed at the 12 icosahedral vertices.[20] Proteins known to form the shell have been structurally characterized by X-ray crystallography. The proteins that constitute the majority of the shell form cyclical hexamers or pseudo-hexamers and belong to the BMC protein family.[21] Small pores perforate many different types of BMC-H hexamers, and may serve as the route for diffusion of small substrates (e.g. bicarbonate) and products (3-phosphoglycerate) into and out of the carboxysome. Positively charged amino acids in the pores presumably help promote the diffusion of the negatively charged substrates and products.[21] Other minor structural components of the shell that have been characterized include pentameric proteins (BMC-P proteins) which occupy the vertices of the icosahedral shell.[22] A third building block of the carboxysome shell is a protein composed of two BMC domains in tandem (BMC-T proteins). Structurally, these are known to form trimers which are pseudohexameric.[23][24] Some members of the BMC-T protein family stack in a face-to-face fashion and form tiny cages, notably both types of carboxysomes (alpha and beta, see below) contain these stacking trimers.[25][26] Based on crystal structures, these protein cages have relatively large gated pores on both sides, and it has been proposed that the opening and closing of the pore could be controlled in a manner similar to an air-lock. Such an air-lock, in contrast to BMC-H proteins with constitutively open pores, has been suggested to serve as a route for larger substrates (ribulose-1,5-bisphosphate) and products (3-phosphoglycerate) that must cross the shell.[23][24]
Production of empty carboxysome shells in E. coli enabled the first visualization of the carboxysome shell by cryo-electron microscopy.[27]
A number of viral capsids are also icosahedral, composed of hexameric and pentameric proteins, but currently there is no evidence suggesting any evolutionary relationship between the carboxysome shell and viral capsids.[28]
Scaffold Proteins
All carboxysomes contain scaffold proteins that nucleate carboxysome components together during the assembly process. These scaffold proteins are required for carboxysome assembly; without them, carboxysomes do not form.[29] The α-carboxysomal scaffold protein is called CsoS2, and the β-carboxysomal scaffold protein is called CcmM. Though CsoS2 and CcmM have related functions, they have no evolutionary or sequence similarity. Both proteins bind to Rubisco, thereby ensuring that Rubisco gets packaged during carboxysome biogenesis.[30][31] Remarkably, both proteins bind to Rubisco at a binding site that bridges two large subunits while maintaining contact with the small subunit, ensuring that only the 16-subunit Rubisco holoenzyme is encapsulated. Both CsoS2 and CcmM have repetitive domain structures giving them multi-valent modes of binding. CcmM has three small-subutnit-like (SSUL) domains that bind to Rubisco,[31] and CsoS2 has four N-terminal domain (NTD) repeats that bind Rubisco,[30] making it possible for each single scaffold protein to bind up to 3-4 Rubiscos at a time. CsoS2 has also been shown to bind to shell proteins via its 7 Middle Region (MR) repeats and C-terminal domain (CTD).[29][32] In α-carboxysomes, the CsoS2 MR repeats have been shown to define the size of the carboxysome.[33]
Two Types of Carboxysomes
There are two types of carboxysomes. Although they may seem similar in appearance, they differ in their protein composition, including the form of RuBisCO they enclose.[34][35][36][37] Furthermore, studies have revealed fundamental differences in their gene organization and possibly their assembly pathway. Based on bioinformatic studies of shell proteins, it appears that the two types of carboxysomes evolved independently.[38][37]
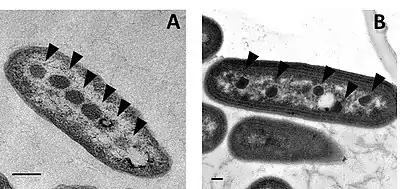
Alpha-Carboxysomes
Alpha-carboxysomes (aka α-carboxysomes) are also referred as the cso type of carboxysome. They contain Form IA RuBisCO; they are found in alpha-cyanobacteria, some nitrifying bacteria, some sulfur-oxidizing bacteria (for example, Halothiobacillus neapolitanus), and some purple bacteria; these are all classified as Pseudomonadota). The alpha-carboxysome was the first bacterial microcompartment to be purified and characterized.[39][40] Electron microscopy studies on purified alpha-carboxysomes or cell sections containing alpha-carboxysomes revealed that they are typically 100-160 nm in diameter.[41] Common building blocks for the shell of alpha-carboxysomes are called CsoS1A/B/C (BMC-H), CsoS4A/B (BMC-P), and CsoS1D (BMC-T). CsoS4A/B were the first BMC-P proteins to be experimentally demonstrated as minor components of the BMC shell[4] (only 12 pentamers are required to cap the vertices of an icosahedron). CsoS1D is the first BMC-T which has been structurally characterized; it is also the first example of dimerization of two BMC building blocks in a face-to-face fashion to create a tiny cage. The CsoS1D cage has a gated pore at both ends, which is proposed to facilitate the transfer of large metabolites across the shell.[24] In addition to the specific form of RuBisCO, other encapsulated proteins distinguish alpha-carboxysomes from beta-carboxysomes such as scaffold protein CsoS2 and carbonic anhydrase CsoSCA. CsoS2 is an intrinsically disordered protein with an essential role in alpha-carboxysome assembly. It has a very high pI and a unique primary structure with three domains: an N-terminal, a middle- and a C-terminal domain.[29][42] Repetitive motifs can be identified in all three regions; the N-terminal domain repeats bind to Rubisco,[30] the middle region domains bind to shell proteins,[32] and the c-terminal domain repeats also bind to shell proteins.[43][44][45] CsoSCA is a beta-carbonic anhydrase that binds to Rubisco.[5][46][47] Studies in Halothiobacillus neapolitanus have shown that empty shells of normal shape and composition are assembled in carboxysomal RuBisCO-lacking mutants, suggesting that alpha-carboxysome shell biogenesis and enzyme sequestration are two independent, but functionally linked processes.[48] Intriguingly, carboxysomes of Halothiobacillus neapolitanus have been found to accommodate chimeric and heterologous species of RuBisCO. It is the large subunit of RuBisCO which determines whether the enzyme is sequestered into carboxysomes.[48]
Beta-carboxysomes
Beta-carboxysomes (aka β-carboxysomes) are found in cyanobacteria.[49]
The signature proteins of the beta-carboxysome are Form IB RuBisCO and a gamma carbonic anhydrase homolog.[9] Beta-carboxysomes are typically larger than alpha-carboxysomes: the observed diameters vary from 200 to 400 nm.[50] The structural proteins that are essential for beta-carboxysome formation are encoded in the conserved carboxysome locus[10] known as the ccm locus. The ccm locus includes genes for core proteins CcmM and CcmN and the shell proteins CcmK (a BMC-H protein), CcmL (a BMC-P protein) and CcmO (a BMC-T protein).
A full length CcmM protein consists of a gamma-carbonic anhydrase domain and three to five RubisCO small subunit-like domains (SSLDs) on its C-terminus.[51] The ccmM gene contains an internal translation site that produces a short form of CcmM which only consists of SSLDs; both long and short forms of CcmM are required for beta-carboxysome assembly.[52] CcmN contains multiple hexapeptide-repeat domains on its N-terminus and a short α-helical encapsulation peptide on the C-terminus.[53]
Other structural components of beta-carboxysomes are encoded outside of the ccm locus. CcmP is a BMC-T protein that is absolutely conserved among organisms that form beta-carboxysomes. Two CcmP pseudohexamers stack to form a nanocompartment—an example of an air-lock forming protein.[23] Likewise, in some cyanobacterial strains the beta-carboxysomes contain a beta-carbonic anhydrase that is not encoded in the ccm locus.[54]
Shell proteins of beta carboxysomes are relatively diverse[49] compared to their counterparts in the alpha carboxysomes, and this has been proposed to reflect variable permeability requirements of beta carboxysomes, which are found in cyanobacteria that occupy ecophysiologically dynamic environments.[55]
The beta-carboxysome assembles from the inside out. First an enzymatic core forms that is subsequently encapsulated by the protein shell.[56] Carboxysome assembly occurs through a series of protein-protein interactions: the enzyme RuBisCO and the two isoforms (full length and short form) of the CcmM protein interact by means of the SSLDs; in strains containing CcaA the beta-carbonic anhydrase is brought into the carboxysome core by interaction with the N-terminus of the full length CcmM.[57][58] Once the procarboxysome (the carboxysome core) is formed, the N-terminus of the adapter protein CcmN interacts with the N-terminus of CcmM, while the C-terminus of CcmN recruits the shell proteins CcmK (BMC-H) and CcmO (BMC-T), utilizing a 15-20 amino acids long peptide.[53] This encapsulation peptide forms an amphipathic a-helix that interacts with the shell components and its role is essential, given that in its absence, carboxysomes cannot be formed.[53][59] The final step is the addition of the vertices formed by the BMC-P protein CcmL, which then cap the enzymatic core and facets.[56] Elucidation of the assembly pathway of beta carboxysomes enabled the design of a single synthetic protein that replaced four other proteins in carboxysome assembly.[60]
Potential uses of the carboxysome in biotechnology
As is the case with other BMCs, the carboxysome is attracting significant attention by researchers for applications in plant synthetic biology.[61][62][63] The transfer of a genetic module coding for an alpha-carboxysome has been shown to produce carboxysome-like structures in E. coli.[64] Bioengineering of carboxysome shells has been shown to be feasible, and beta-carboxysomes constructed with chimeric proteins or with chimeric shells have been reported.[65] The introduction of carboxysomes into plant chloroplasts as part of a CO2 concentrating mechanism [66][67] such as that found in cyanobacteria is predicted to significantly improve net CO2 fixation and yield.[68][69] Expression of beta-carboxysomal shell proteins [70] and Form IB Rubisco-CcmM complexes in tobacco chloroplasts has been achieved,[71] but did not result in compartments containing RuBisCO. A further advance has been the construction of minimal alpha-carboxysomes containing Form IA Rubisco and the CsoS1A and CsoS2 proteins from the cyanobacterium Cyanobium PCC7001 in tobacco chloroplasts.[72] As yet, identifiably functional carboxysomes have not been constructed in plant chloroplasts. Improvement of photosynthesis in plants using this approach is ultimately dependent on the operation of transporter proteins in the chloroplast inner envelope membrane to help generate a high concentration of bicarbonate inside the chloroplast.[73]
Potential applications of carboxysomes (list format):
- Engineer the carbon-concentrating mechanism (CCM) and carboxysomes into industrially relevant microbes, potentially converting heterotrophic organisms into mixotrophs or autotrophs that capture CO2 while producing high value products.[74]
- Engineer the carbon-concentrating mechanism (CCM) and carboxysomes into plants for increased CO2 capture and enhanced growth.
- Engineer faster Rubiscos. The fastest form I prokaryotic Rubiscos are mostly found in α-carboxysomes.[75]
- Engineer a minimal carboxysome gene set (Rubisco, carbonic anhydrase, scaffold protein, hexameric shell, pentameric shell) to facilitate facile engineering into alternative host organisms.
- Design in vitro carboxysomes for cell-free CO2 fixation.
- Engineer carboxysomes to have alternative metabolisms.[44][76]
Carboxysome Reviews (by year)
Carboxysome research expands every year. Published reviews chart the rapid pace of discovery across the broad field of "carboxysomics".
References
- Tsai Y, Sawaya MR, Cannon GC, Cai F, Williams EB, Heinhorst S, et al. (June 2007). "Structural analysis of CsoS1A and the protein shell of the Halothiobacillus neapolitanus carboxysome". PLOS Biology. 5 (6): e144. doi:10.1371/journal.pbio.0050144. PMC 1872035. PMID 17518518.
- Yeates TO, Kerfeld CA, Heinhorst S, Cannon GC, Shively JM (September 2008). "Protein-based organelles in bacteria: carboxysomes and related microcompartments". Nature Reviews. Microbiology. 6 (9): 681–691. doi:10.1038/nrmicro1913. PMID 18679172. S2CID 22666203.
- Badger MR, Price GD (February 2003). "CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution". Journal of Experimental Botany. 54 (383): 609–622. doi:10.1093/jxb/erg076. PMID 12554704.
- Cai F, Menon BB, Cannon GC, Curry KJ, Shively JM, Heinhorst S (October 2009). "The pentameric vertex proteins are necessary for the icosahedral carboxysome shell to function as a CO2 leakage barrier". PLOS ONE. 4 (10): e7521. Bibcode:2009PLoSO...4.7521C. doi:10.1371/journal.pone.0007521. PMC 2760150. PMID 19844578.
- Dou Z, Heinhorst S, Williams EB, Murin CD, Shively JM, Cannon GC (April 2008). "CO2 fixation kinetics of Halothiobacillus neapolitanus mutant carboxysomes lacking carbonic anhydrase suggest the shell acts as a diffusional barrier for CO2". The Journal of Biological Chemistry. 283 (16): 10377–10384. doi:10.1074/jbc.M709285200. PMID 18258595.
- Mangan NM, Flamholz A, Hood RD, Milo R, Savage DF (September 2016). "pH determines the energetic efficiency of the cyanobacterial CO2 concentrating mechanism". Proceedings of the National Academy of Sciences of the United States of America. 113 (36): E5354–E5362. Bibcode:2016PNAS..113E5354M. doi:10.1073/pnas.1525145113. PMC 5018799. PMID 27551079.
- Badger MR, Hanson D, Price GD (April 2002). "Evolution and diversity of CO2 concentrating mechanisms in cyanobacteria". Functional Plant Biology. 29 (3): 161–173. doi:10.1071/PP01213. PMID 32689463.
- Price GD (September 2011). "Inorganic carbon transporters of the cyanobacterial CO2 concentrating mechanism". Photosynthesis Research. 109 (1–3): 47–57. doi:10.1007/s11120-010-9608-y. PMID 21359551. S2CID 25867128.
- Kerfeld CA, Erbilgin O (January 2015). "Bacterial microcompartments and the modular construction of microbial metabolism". Trends in Microbiology. 23 (1): 22–34. doi:10.1016/j.tim.2014.10.003. PMID 25455419.
- Axen SD, Erbilgin O, Kerfeld CA (October 2014). "A taxonomy of bacterial microcompartment loci constructed by a novel scoring method". PLOS Computational Biology. 10 (10): e1003898. Bibcode:2014PLSCB..10E3898A. doi:10.1371/journal.pcbi.1003898. PMC 4207490. PMID 25340524.
- Drews G, Niklowitz W (1956). "[Cytology of Cyanophycea. II. Centroplasm and granular inclusions of Phormidium uncinatum]". Archiv Fur Mikrobiologie. 24 (2): 147–162. doi:10.1007/BF00408629. PMID 13327992. S2CID 46171409.
- Gantt E, Conti SF (March 1969). "Ultrastructure of blue-green algae". Journal of Bacteriology. 97 (3): 1486–1493. doi:10.1128/JB.97.3.1486-1493.1969. PMC 249872. PMID 5776533.
- Shively JM (1974). "Inclusion bodies of prokaryotes". Annual Review of Microbiology. 28: 167–187. doi:10.1146/annurev.mi.28.100174.001123. PMID 4372937.
- Shively JM, Ball F, Brown DH, Saunders RE (November 1973). "Functional organelles in prokaryotes: polyhedral inclusions (carboxysomes) of Thiobacillus neapolitanus". Science. 182 (4112): 584–586. Bibcode:1973Sci...182..584S. doi:10.1126/science.182.4112.584. PMID 4355679. S2CID 10097616.
- Iancu CV, Ding HJ, Morris DM, Dias DP, Gonzales AD, Martino A, Jensen GJ (September 2007). "The structure of isolated Synechococcus strain WH8102 carboxysomes as revealed by electron cryotomography". Journal of Molecular Biology. 372 (3): 764–773. doi:10.1016/j.jmb.2007.06.059. PMC 2453779. PMID 17669419.
- Iancu CV, Morris DM, Dou Z, Heinhorst S, Cannon GC, Jensen GJ (February 2010). "Organization, structure, and assembly of alpha-carboxysomes determined by electron cryotomography of intact cells". Journal of Molecular Biology. 396 (1): 105–117. doi:10.1016/j.jmb.2009.11.019. PMC 2853366. PMID 19925807.
- Schmid MF, Paredes AM, Khant HA, Soyer F, Aldrich HC, Chiu W, Shively JM (December 2006). "Structure of Halothiobacillus neapolitanus carboxysomes by cryo-electron tomography". Journal of Molecular Biology. 364 (3): 526–535. doi:10.1016/j.jmb.2006.09.024. hdl:11147/2128. PMC 1839851. PMID 17028023.
- Metskas LA, Ortega D, Oltrogge LM, Blikstad C, Lovejoy DR, Laughlin TG, et al. (August 2022). "Rubisco forms a lattice inside alpha-carboxysomes". Nature Communications. 13 (1): 4863. Bibcode:2022NatCo..13.4863M. doi:10.1038/s41467-022-32584-7. PMC 9388693. PMID 35982043.
- Vernizzi G, Sknepnek R, Olvera de la Cruz M (March 2011). "Platonic and Archimedean geometries in multicomponent elastic membranes". Proceedings of the National Academy of Sciences of the United States of America. 108 (11): 4292–4296. doi:10.1073/pnas.1012872108. PMC 3060260. PMID 21368184.
- Kinney JN, Axen SD, Kerfeld CA (September 2011). "Comparative analysis of carboxysome shell proteins". Photosynthesis Research. 109 (1–3): 21–32. doi:10.1007/s11120-011-9624-6. PMC 3173617. PMID 21279737.
- Kerfeld CA, Sawaya MR, Tanaka S, Nguyen CV, Phillips M, Beeby M, Yeates TO (August 2005). "Protein structures forming the shell of primitive bacterial organelles". Science. 309 (5736): 936–938. Bibcode:2005Sci...309..936K. CiteSeerX 10.1.1.1026.896. doi:10.1126/science.1113397. PMID 16081736. S2CID 24561197.
- Tanaka S, Kerfeld CA, Sawaya MR, Cai F, Heinhorst S, Cannon GC, Yeates TO (February 2008). "Atomic-level models of the bacterial carboxysome shell". Science. 319 (5866): 1083–1086. Bibcode:2008Sci...319.1083T. doi:10.1126/science.1151458. PMID 18292340. S2CID 5734731.
- Cai F, Sutter M, Cameron JC, Stanley DN, Kinney JN, Kerfeld CA (May 2013). "The structure of CcmP, a tandem bacterial microcompartment domain protein from the β-carboxysome, forms a subcompartment within a microcompartment". The Journal of Biological Chemistry. 288 (22): 16055–16063. doi:10.1074/jbc.M113.456897. PMC 3668761. PMID 23572529.
- Klein MG, Zwart P, Bagby SC, Cai F, Chisholm SW, Heinhorst S, et al. (September 2009). "Identification and structural analysis of a novel carboxysome shell protein with implications for metabolite transport". Journal of Molecular Biology. 392 (2): 319–333. doi:10.1016/j.jmb.2009.03.056. hdl:1721.1/61355. PMID 19328811. S2CID 42771660.
- Cai F, Sutter M, Cameron JC, Stanley DN, Kinney JN, Kerfeld CA (May 2013). "The structure of CcmP, a tandem bacterial microcompartment domain protein from the β-carboxysome, forms a subcompartment within a microcompartment". The Journal of Biological Chemistry. 288 (22): 16055–16063. doi:10.1074/jbc.M113.456897. PMC 3668761. PMID 23572529.
- Klein MG, Zwart P, Bagby SC, Cai F, Chisholm SW, Heinhorst S, et al. (September 2009). "Identification and structural analysis of a novel carboxysome shell protein with implications for metabolite transport". Journal of Molecular Biology. 392 (2): 319–333. doi:10.1016/j.jmb.2009.03.056. hdl:1721.1/61355. PMID 19328811. S2CID 42771660.
- Sutter M, Laughlin TG, Sloan NB, Serwas D, Davies KM, Kerfeld CA (November 2019). "Structure of a Synthetic β-Carboxysome Shell". Plant Physiology. 181 (3): 1050–1058. doi:10.2210/pdb6owg/pdbx. PMC 6836842. PMID 31501298.
- Krupovic M, Koonin EV (November 2017). "Cellular origin of the viral capsid-like bacterial microcompartments". Biology Direct. 12 (1): 25. doi:10.1186/s13062-017-0197-y. PMC 5683377. PMID 29132422.
- Cai F, Dou Z, Bernstein SL, Leverenz R, Williams EB, Heinhorst S, et al. (March 2015). "Advances in Understanding Carboxysome Assembly in Prochlorococcus and Synechococcus Implicate CsoS2 as a Critical Component". Life. 5 (2): 1141–1171. Bibcode:2015Life....5.1141C. doi:10.3390/life5021141. PMC 4499774. PMID 25826651.
- Oltrogge LM, Chaijarasphong T, Chen AW, Bolin ER, Marqusee S, Savage DF (March 2020). "Multivalent interactions between CsoS2 and Rubisco mediate α-carboxysome formation". Nature Structural & Molecular Biology. 27 (3): 281–287. doi:10.1038/s41594-020-0387-7. PMC 7337323. PMID 32123388.
- Wang H, Yan X, Aigner H, Bracher A, Nguyen ND, Hee WY, et al. (February 2019). "Rubisco condensate formation by CcmM in β-carboxysome biogenesis". Nature. 566 (7742): 131–135. Bibcode:2019Natur.566..131W. doi:10.1038/s41586-019-0880-5. hdl:1885/173112. PMID 30675061. S2CID 256770108.
- Turnšek JB, Oltrogge LM, Savage DF (2023-07-08). "Conserved and repetitive motifs in an intrinsically disordered protein drive α-carboxysome assembly". bioRxiv. Cold Spring Harbor Laboratory (CSHL). doi:10.1101/2023.07.08.548221. S2CID 259834050.
- Oltrogge LM, Chen AW, Chaijarasphong T, Turnšek JB, Savage DF (2023-07-08). "α-carboxysome size is controlled by the disordered scaffold protein CsoS2". bioRxiv. Cold Spring Harbor Laboratory (CSHL). doi:10.1101/2023.07.07.548173. S2CID 259834078.
- Zarzycki J, Axen SD, Kinney JN, Kerfeld CA (January 2013). "Cyanobacterial-based approaches to improving photosynthesis in plants". Journal of Experimental Botany. 64 (3): 787–798. doi:10.1093/jxb/ers294. PMID 23095996.
- Rae BD, Long BM, Badger MR, Price GD (September 2013). "Functions, compositions, and evolution of the two types of carboxysomes: polyhedral microcompartments that facilitate CO2 fixation in cyanobacteria and some proteobacteria". Microbiology and Molecular Biology Reviews. 77 (3): 357–379. doi:10.1128/MMBR.00061-12. PMC 3811607. PMID 24006469.
- Turmo A, Gonzalez-Esquer CR, Kerfeld CA (October 2017). "Carboxysomes: metabolic modules for CO2 fixation". FEMS Microbiology Letters. 364 (18). doi:10.1093/femsle/fnx176. PMID 28934381.
- Kerfeld CA, Melnicki MR (June 2016). "Assembly, function and evolution of cyanobacterial carboxysomes". Current Opinion in Plant Biology. 31: 66–75. doi:10.1016/j.pbi.2016.03.009. PMID 27060669.
- Melnicki MR, Sutter M, Kerfeld CA (October 2021). "Evolutionary relationships among shell proteins of carboxysomes and metabolosomes". Current Opinion in Microbiology. 63: 1–9. doi:10.1016/j.mib.2021.05.011. PMC 8525121. PMID 34098411.
- Shively JM, Bock E, Westphal K, Cannon GC (November 1977). "Icosahedral inclusions (carboxysomes) of Nitrobacter agilis". Journal of Bacteriology. 132 (2): 673–675. doi:10.1128/JB.132.2.673-675.1977. PMC 221910. PMID 199579.
- Cannon GC, Shively JM (1983). "Characterization of a homogenous preparation of carboxysomes from Thiobacillus neapolitanus". Archives of Microbiology. 134 (1): 52–59. doi:10.1007/BF00429407. ISSN 0302-8933. S2CID 22329896.
- Heinhorst S, Cannon GC, Shively JM (2014). "Carboxysomes and Their Structural Organization in Prokaryotes". Nanomicrobiology. pp. 75–101. doi:10.1007/978-1-4939-1667-2_4. ISBN 978-1-4939-1666-5.
- Borden JS, Savage DF (June 2021). "New discoveries expand possibilities for carboxysome engineering". Current Opinion in Microbiology. 61: 58–66. doi:10.1016/j.mib.2021.03.002. PMC 8169590. PMID 33798818.
- Tan YQ, Ali S, Xue B, Teo WZ, Ling LH, Go MK, et al. (October 2021). "Structure of a Minimal α-Carboxysome-Derived Shell and Its Utility in Enzyme Stabilization". Biomacromolecules. 22 (10): 4095–4109. doi:10.1021/acs.biomac.1c00533. PMID 34384019. S2CID 236999059.
- Li T, Jiang Q, Huang J, Aitchison CM, Huang F, Yang M, et al. (October 2020). "Reprogramming bacterial protein organelles as a nanoreactor for hydrogen production". Nature Communications. 11 (1): 5448. Bibcode:2020NatCo..11.5448L. doi:10.1038/s41467-020-19280-0. PMC 7595155. PMID 33116131.
- Ni T, Jiang Q, Ng PC, Shen J, Dou H, Zhu Y, et al. (2023). "Intrinsically disordered CsoS2 acts as a general molecular thread for α-carboxysome shell assembly". bioRxiv. Cold Spring Harbor Laboratory (CSHL). doi:10.1101/2023.06.24.546370. S2CID 259276395.
- Sawaya MR, Cannon GC, Heinhorst S, Tanaka S, Williams EB, Yeates TO, Kerfeld CA (March 2006). "The structure of beta-carbonic anhydrase from the carboxysomal shell reveals a distinct subclass with one active site for the price of two". The Journal of Biological Chemistry. 281 (11): 7546–7555. doi:10.1074/jbc.M510464200. PMID 16407248.
- Blikstad C, Dugan EJ, Laughlin TG, Turnšek JB, Liu MD, Shoemaker SR, et al. (2023). "Discovery of a carbonic anhydrase-Rubisco complex within the alpha-carboxysome". bioRxiv. Cold Spring Harbor Laboratory (CSHL). doi:10.1101/2021.11.05.467472. S2CID 243836266.
- Menon BB, Dou Z, Heinhorst S, Shively JM, Cannon GC (2008). "Halothiobacillus neapolitanus carboxysomes sequester heterologous and chimeric RubisCO species". PLOS ONE. 3 (10): e3570. Bibcode:2008PLoSO...3.3570M. doi:10.1371/journal.pone.0003570. PMC 2570492. PMID 18974784.
- Sommer M, Cai F, Melnicki M, Kerfeld CA (June 2017). "β-Carboxysome bioinformatics: identification and evolution of new bacterial microcompartment protein gene classes and core locus constraints". Journal of Experimental Botany. 68 (14): 3841–3855. doi:10.1093/jxb/erx115. PMC 5853843. PMID 28419380.
- Cai F, Dou Z, Bernstein SL, Leverenz R, Williams EB, Heinhorst S, et al. (March 2015). "Advances in Understanding Carboxysome Assembly in Prochlorococcus and Synechococcus Implicate CsoS2 as a Critical Component". Life. 5 (2): 1141–1171. Bibcode:2015Life....5.1141C. doi:10.3390/life5021141. PMC 4499774. PMID 25826651.
- Long BM, Badger MR, Whitney SM, Price GD (October 2007). "Analysis of carboxysomes from Synechococcus PCC7942 reveals multiple Rubisco complexes with carboxysomal proteins CcmM and CcaA". The Journal of Biological Chemistry. 282 (40): 29323–29335. doi:10.1074/jbc.M703896200. PMID 17675289.
- Long BM, Tucker L, Badger MR, Price GD (May 2010). "Functional cyanobacterial beta-carboxysomes have an absolute requirement for both long and short forms of the CcmM protein". Plant Physiology. 153 (1): 285–293. doi:10.1104/pp.110.154948. PMC 2862411. PMID 20304968.
- Kinney JN, Salmeen A, Cai F, Kerfeld CA (May 2012). "Elucidating essential role of conserved carboxysomal protein CcmN reveals common feature of bacterial microcompartment assembly". The Journal of Biological Chemistry. 287 (21): 17729–17736. doi:10.1074/jbc.M112.355305. PMC 3366800. PMID 22461622.
- Cannon GC, Heinhorst S, Kerfeld CA (February 2010). "Carboxysomal carbonic anhydrases: Structure and role in microbial CO2 fixation". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1804 (2): 382–392. doi:10.1016/j.bbapap.2009.09.026. PMID 19818881. S2CID 3712407.
- Sommer M, Sutter M, Gupta S, Kirst H, Turmo A, Lechno-Yossef S, et al. (January 2019). "Heterohexamers Formed by CcmK3 and CcmK4 Increase the Complexity of Beta Carboxysome Shells". Plant Physiology. 179 (1): 156–167. doi:10.1104/pp.18.01190. PMC 6324227. PMID 30389783.
- Cameron JC, Wilson SC, Bernstein SL, Kerfeld CA (November 2013). "Biogenesis of a bacterial organelle: the carboxysome assembly pathway". Cell. 155 (5): 1131–1140. doi:10.1016/j.cell.2013.10.044. PMID 24267892.
- Cot SS, So AK, Espie GS (February 2008). "A multiprotein bicarbonate dehydration complex essential to carboxysome function in cyanobacteria". Journal of Bacteriology. 190 (3): 936–945. doi:10.1128/JB.01283-07. PMC 2223583. PMID 17993516.
- Long BM, Rae BD, Badger MR, Price GD (September 2011). "Over-expression of the β-carboxysomal CcmM protein in Synechococcus PCC7942 reveals a tight co-regulation of carboxysomal carbonic anhydrase (CcaA) and M58 content". Photosynthesis Research. 109 (1–3): 33–45. doi:10.1007/s11120-011-9659-8. PMID 21597987. S2CID 20716799.
- Kerfeld CA, Melnicki MR (June 2016). "Assembly, function and evolution of cyanobacterial carboxysomes". Current Opinion in Plant Biology. 31: 66–75. doi:10.1016/j.pbi.2016.03.009. PMID 27060669.
- Gonzalez-Esquer CR, Shubitowski TB, Kerfeld CA (September 2015). "Streamlined Construction of the Cyanobacterial CO2-Fixing Organelle via Protein Domain Fusions for Use in Plant Synthetic Biology". The Plant Cell. 27 (9): 2637–2644. doi:10.1105/tpc.15.00329. PMC 4815102. PMID 26320224.
- Kerfeld CA (December 2015). "Plug-and-play for improving primary productivity". American Journal of Botany. 102 (12): 1949–1950. doi:10.3732/ajb.1500409. PMID 26656128.
- Zarzycki J, Axen SD, Kinney JN, Kerfeld CA (January 2013). "Cyanobacterial-based approaches to improving photosynthesis in plants". Journal of Experimental Botany. 64 (3): 787–798. doi:10.1093/jxb/ers294. PMID 23095996.
- Gonzalez-Esquer CR, Newnham SE, Kerfeld CA (July 2016). "Bacterial microcompartments as metabolic modules for plant synthetic biology". The Plant Journal. 87 (1): 66–75. doi:10.1111/tpj.13166. PMID 26991644.
- Bonacci W, Teng PK, Afonso B, Niederholtmeyer H, Grob P, Silver PA, Savage DF (January 2012). "Modularity of a carbon-fixing protein organelle". Proceedings of the National Academy of Sciences of the United States of America. 109 (2): 478–483. doi:10.1073/pnas.1108557109. PMC 3258634. PMID 22184212.
- Cai F, Sutter M, Bernstein SL, Kinney JN, Kerfeld CA (April 2015). "Engineering bacterial microcompartment shells: chimeric shell proteins and chimeric carboxysome shells". ACS Synthetic Biology. 4 (4): 444–453. doi:10.1021/sb500226j. PMID 25117559.
- Price GD, Badger MR, Woodger FJ, Long BM (2008). "Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants". Journal of Experimental Botany. 59 (7): 1441–1461. doi:10.1093/jxb/erm112. PMID 17578868.
- Price GD, Pengelly JJ, Forster B, Du J, Whitney SM, von Caemmerer S, et al. (January 2013). "The cyanobacterial CCM as a source of genes for improving photosynthetic CO2 fixation in crop species". Journal of Experimental Botany. 64 (3): 753–768. doi:10.1093/jxb/ers257. hdl:1885/53387. PMID 23028015.
- McGrath JM, Long SP (April 2014). "Can the cyanobacterial carbon-concentrating mechanism increase photosynthesis in crop species? A theoretical analysis". Plant Physiology. 164 (4): 2247–2261. doi:10.1104/pp.113.232611. PMC 3982776. PMID 24550242.
- Yin X, Struik PC (April 2017). "Can increased leaf photosynthesis be converted into higher crop mass production? A simulation study for rice using the crop model GECROS". Journal of Experimental Botany. 68 (9): 2345–2360. doi:10.1093/jxb/erx085. PMC 5447886. PMID 28379522.
- Lin MT, Occhialini A, Andralojc PJ, Devonshire J, Hines KM, Parry MA, Hanson MR (July 2014). "β-Carboxysomal proteins assemble into highly organized structures in Nicotiana chloroplasts". The Plant Journal. 79 (1): 1–12. doi:10.1111/tpj.12536. PMC 4080790. PMID 24810513.
- Lin MT, Occhialini A, Andralojc PJ, Parry MA, Hanson MR (September 2014). "A faster Rubisco with potential to increase photosynthesis in crops". Nature. 513 (7519): 547–550. Bibcode:2014Natur.513..547L. doi:10.1038/nature13776. PMC 4176977. PMID 25231869.
- Long BM, Hee WY, Sharwood RE, Rae BD, Kaines S, Lim YL, et al. (September 2018). "Carboxysome encapsulation of the CO2-fixing enzyme Rubisco in tobacco chloroplasts". Nature Communications. 9 (1): 3570. Bibcode:2018NatCo...9.3570L. doi:10.1038/s41467-018-06044-0. PMC 6120970. PMID 30177711.
- Rae BD, Long BM, Förster B, Nguyen ND, Velanis CN, Atkinson N, et al. (June 2017). "Progress and challenges of engineering a biophysical CO2-concentrating mechanism into higher plants". Journal of Experimental Botany. 68 (14): 3717–3737. doi:10.1093/jxb/erx133. PMID 28444330..
- Flamholz AI, Dugan E, Blikstad C, Gleizer S, Ben-Nissan R, Amram S, et al. (October 2020). "Functional reconstitution of a bacterial CO2 concentrating mechanism in Escherichia coli". eLife. 9. doi:10.7554/eLife.59882. PMC 7714395. PMID 33084575.
- de Pins B, Greenspoon L, Bar-On YM, Shamshoum M, Ben-Nissan R, Milshtein E, et al. (2023-07-27). "Systematic exploration of prokaryotic form I rubisco maximal carboxylation rates". bioRxiv. Cold Spring Harbor Laboratory (CSHL). doi:10.1101/2023.07.27.550689. S2CID 260289274.
- Jiang Q, Li T, Yang J, Aitchison CM, Huang J, Chen Y, et al. (March 2023). "Synthetic engineering of a new biocatalyst encapsulating [NiFe]-hydrogenases for enhanced hydrogen production". Journal of Materials Chemistry. B. 11 (12): 2684–2692. doi:10.1039/D2TB02781J. PMC 10032307. PMID 36883480.