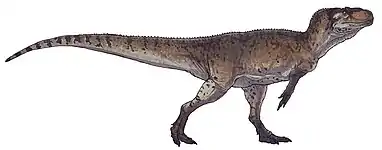
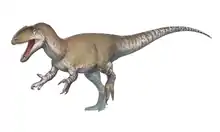Carcharodontosaurus
Carcharodontosaurus (/ˌkɑːrkəroʊˌdɒntoʊˈsɔːrəs/; lit. 'shark-toothed lizard') is a genus of carnivorous theropod dinosaur that lived in North Africa from about 99 to 94 million years ago during the Albian and Cenomanian stages of the Late Cretaceous. Two teeth of the genus, now lost, were first described from Algeria by French paleontologists Charles Depéret and Justin Savornin as Megalosaurus saharicus. A partial skeleton was collected by crews of German paleontologist Ernst Stromer during a 1914 expedition to Egypt. Stromer did not report the Egyptian find until 1931, in which he dubbed the novel genus Carcharodontosaurus, making the type species C. saharicus. Unfortunately, this skeleton was destroyed during the Second World War. In 1995 a nearly complete skull of C. saharicus, the first well-preserved specimen to be found in almost a century, was discovered in the Kem Kem Beds of Morocco; it was designated the neotype in 1996. Fossils unearthed from the Echkar Formation of northern Niger were described and named as another species, C. iguidensis, in 2007.
| Carcharodontosaurus Temporal range: Late Cretaceous (Cenomanian), | |
|---|---|
 | |
| Reconstructed C. saharicus skull, Science Museum of Minnesota | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Carcharodontosauridae |
| Subfamily: | †Carcharodontosaurinae |
| Genus: | †Carcharodontosaurus Stromer, 1931 |
| Type species | |
| †Carcharodontosaurus saharicus Depéret & Savornin, 1925 | |
| Other species | |
| Synonyms | |
|
List
| |
Carcharodontosaurus is one of the largest theropod dinosaurs known, with C. saharicus reaching 12–12.5 metres (39–41 ft) in length and approximately 6–6.2 metric tons (6.6–6.8 short tons) in body mass. It had a large, lightly built skull with a triangular rostrum. Its jaws were lined with sharp, recurved, serrated teeth that bear striking resemblances to those of the great white shark, the inspiration for the name. Though giant, its cranium was made lighter by greatly expanded fossae and fenestra, but also making it more fragile than tyrannosaurids'. The forelimbs were tiny whereas the hindlimbs were robust and muscular. Like most other theropods, it had an elongated tail for balance.
Many gigantic theropods are known from North Africa during this period, including both species of Carcharodontosaurus as well as the spinosaurid Spinosaurus and ceratosaur Deltadromeus. This suggests there was niche partitioning between the different groups, with Spinosaurus being piscivorous whereas Carcharodontosaurus consumed sauropod dinosaurs. Studies of the bite force and tooth anatomy of carcharodontosaurids have found them to possess strong, but relatively weak bites compared to other theropod families. North Africa at the time was blanketed in mangrove forests and wetlands, creating a hotspot of fish, crocodilian, and pterosaur diversity.
Discovery and species
Initial finds
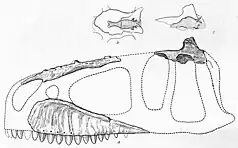
In 1924, two teeth of Carcharodontosaurus were unearthed from wall cuts in different foggaras near Timimoun, French Algeria. These sedimens came from the Cretaceous-aged[1] Continental intercalaire Formation.[2] The fossils were taken to the governor of Timimoun, Captain Burté, who gave them to French geologist Charles Depéret later that year. In 1925, Depéret and his colleague Justin Savornin described the teeth as syntypes (name-bearing specimens) of a new species of theropod dinosaur, Megalosaurus saharicus. These were the first fossils of theropods to be described from the region.[3] The name saharicus refers to the Sahara Desert where the teeth had been found.[4] The genus Megalosaurus was a wastebasket taxon, with many new species referred to it without justification, including M. saharicus.[5] It was later considered to be a species of Dryptosaurus in 1927,[2] though this is unjustified.[6][7] By accident, another species of Megalosaurus, M. africanus, was named by German paleontologist Friedrich von Huene based on the teeth.[6] It is therefore considered a junior synonym of M. saharicus. Both syntypic teeth of M. saharicus have since been lost, possibly being kept in a collection in Algeria, Paris, or Lyon, and lack distinguishing characteristics from other carcharodontosaurids.[8] In 1960, French paleontologist Albert-Félix de Lapparent reported the discovery of more teeth and several caudal vertebrae from sites in Algeria belonging to Carcharodontosaurus,[9] though some of these fossils might belong to other genera.[10] Later authors have mentioned finds of teeth and isolated fossils from other provinces of Algeria.[11][1]
However, fossils of C. saharicus were first found in marls near Ain Gedid, Egypt in early April 1914 by Austro-Hungarian paleontologist Richard Markgraf. Marls from this region derive from the Cenomanian-aged Bahariya Formation, one of many Cretaceous-aged sites of North Africa.[7][12][13] In Bahariya, Markgraf did extensive collecting of dinosaur skeletons for his employer, German paleontologist Ernst Stromer of the Paläontologisches Museum München (Bavarian State Collection of Paleontology). The skeleton of Carcharodontosaurus (IPHG 1922 X46) consisted of: a partial skull, including much of the braincase, teeth, three cervical and a caudal vertebra, incomplete pelvis, a manual ungual, femora, and the left fibula. An isolated ilium was also considered to be from C. saharicus,[14] but it is likely from a ceratosaur.[10]
Due to political tensions between the German Empire and then British-owned Egypt, the Carcharodontosaurus skeleton took years to get to Germany. It was not until 1922 that they were transported overseas to Munich where they were described by Stromer in 1931.[15] Stromer recognized that IPHG 1922 X46’s teeth matched the characteristic dentition of those described by Depéret and Savornin, which led to Stromer conserving the species name saharicus. However, he found it necessary to erect a new genus for this species, Carcharodontosaurus, for their similarities, in sharpness and serrations, to the teeth of the great white shark (Carcharodon carcharias).[13] World War II would break out in 1939, leading IPHG 1922 X46 and other material from Bahariya to be destroyed during a British bombing raid on Munich during the night of April 24/25, 1944.[16][17] An endocast was made and survived the war, being the only remaining relic of the specimen.[12]
Resurgent discoveries, C. iguidensis, and confusion with spinosaurids
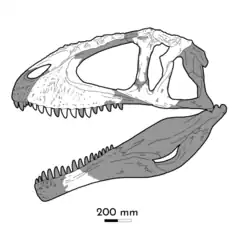
Few discoveries of Carcharodontosaurus material were made until 1995, when American paleontologist Paul Sereno found an incomplete skull during an expedition embarked on by the University of Chicago. This skull (UCRC PV12) was found in the Cenomanian-aged rocks of the Lower Douira Formation, Kem Kem Beds in Errachidia, southeastern Morocco. The specimen was taken to the University of Chicago and described in 1996 by Sereno and colleagues. In a later paper, UCRC PV12 was designated as the neotype of C. saharicus due to the loss of other specimens and the similar age and geographic location to previously noted material. The taxonomy of Carcharodontosaurus was discussed by Chiarenza and Cau (2016),[18] who suggested that the neotype of C. saharicus was similar but distinct from the holotype in the morphology of the maxillary interdental plates. However, paleontologist Mickey Mortimer put forward that the suggested difference between the C. saharicus neotype and holotype was actually due to damage to the neotype.[10]
Several other fossils of C. saharicus have been unearthed from the Kem Kem Beds, such as dentary fragments, a cervical vertebra, and many teeth.[19][20][12] Sereno et al also referred a multitude of cervical vertebrae described as the spinosaurids Sigilmassasaurus and "Spinosaurus B" to C. saharicus reasoning that stout cervicals would be needed to carry the skulls of carcharodontosaurids.[8][7] Later research proved otherwise, with the vertebrae being placed in Spinosaurus aegyptiacus by Ibrahim et al (2020).[12] French paleontologist René Lavocat was the first to note the possible presence of Carcharodontosaurus in Morocco as early as 1954.[21]
In 2007, a novel species of Carcharodontosaurus, C. iguidensis, was dubbed by paleontologists Steve Brusatte and Paul Sereno. Fossils of C. iguidensis had been uncovered during an expedition to the Echkar Formation of Iguidi, Niger, a partial maxilla (MNN IGU2) being designated the holotype. The species name iguidensis is after Iguidi, where the fossils were unearthed. Several other remains such as a braincase, a lacrimal, a dentary, a cervical vertebra, and a collection of teeth were referred to C. iguidensis based on size and supposed similarities to other Carcharodontosaurus bones. Chiarenza and Cau (2016) identified the referred material of C. iguidensis as belonging to Sigilmassasaurus (later referred to Spinosaurus sp.)[22] and a non-carcharodontosaurine, and therefore chose to limit C. iguidensis to the holotype pending future research.[18] Another carcharodontosaurid from the Kem Kem Beds, Sauroniops pachytholus, was dubbed in 2012 based on a single frontal.[23] This species has been proposed to be synonymous with C. saharicus,[12] though this has seen resistance and the validity maintained in most literature.[23][24][18][25] The South American genus Giganotosaurus was synonymized with Carcharodontosaurus by Figueiredo (1998) and Paul (2010),[26][27] however no authors have since followed this assessment.[18]
Other referred specimens
- Lapparent (1951, 1960) described several Carcharodontosaurus teeth from the Continental intercalaire Formation of Guermessa, Tunisia.[3]
- A postorbital bone and several postcranial remains assigned to Carcharodontosaurus were found in the Elrhaz Formation of northern Niger. Taquet (1976) noted that the postorbital was similar to that of Acrocanthosaurus, a relative of Carcharodontosaurus,[28] while the postcranial fossils could belong to other theropods.[10]
- Two braincase fragments, 137 teeth, two caudal vertebrae, and a manual phalanx from the Echkar Formation were referred to as Carcharodontosaurus by Lapparent (1960).[9] A pedal phalanx had also been described as Carcharodontosaurus but it likely is from a spinosaurid instead.[29]
- Many vertebrae, including two associated dorsals, were found in the Early Cretaceous strata of the Irhazer Group of Agadez, Niger. Lapparent mentioned these fossils as C. saharicus in 1960,[9] though they may belong to other theropod genera.[10]
- Caudal vertebrae from the Tefidet and teeth from Akarazeras sites of the Continental intercalaire Formation of Agadez, Niger were recorded by Lapparent (1960)[9] and Taquet (1976) respectively.[28] The vertebrae could be from other theropods.[10]
- From an unknown locale in the Continental intercalaire of the Sahara Desert, Lapparent (1960) documented eight vertebrae, a humerus, and a manual phalanx as coming from C. saharicus.[9] These elements could be from other theropods.[10]
- Two papers, the first in 1978, have described teeth and a caudal vertebra of Carcharodontosaurus from the Chenini Formation of southern Tunisia.[30] However, the caudal vertebra is now labeled Carcharodontosauridae indet.[31]
- Fossils from the Quseir Formation of western Egypt were assigned to Carcharodontosaurus in 1999.[32]
- A 1999 paper described several vertebrae and limb elements of a carcharodontosaurid which were unearthed from the Wadi Milk Formation of Sudan. These were placed only as carcharodontosaurid, but researcher Mickey Mortimer proposed that they belong to Carcharodontosaurus itself based on the presence of pleurocoels (shallow openings) in the caudal vertebrae.[10]
- In 2015, a large neural arch of a dorsal vertebra from the Kem Kem Beds was informally described as belonging to a new genus and species of megaraptoran dubbed "Osteoporosia gigantea". This specimen is owned head of a Polish theme park chain who described it as belonging to a 15 metres (49 ft) long carnosaur similar to Mapusaurus and Carcharodontosaurus.[33] However, it was much smaller than proposed and may belong to C. saharicus or Sauroniops based on its carcharodontosaurid traits and origin.[34]
Erroneously assigned specimens
- A maxillary tooth recovered from the Villar del Arzobispo Formation of Spain was referred to Carcharodontosaurus in 1966.[35] However, it lacks the traits of carcharodontosaurid teeth and instead is more similar to that of other allosauroids.[36][37]
- Bond and Bromley (1970) described teeth deriving from the Gokwe Formation of Zimbabwe as being similar to Carcharodontosaurus, with Mickey Mortimer assigning them to the genus tentatively.[38] However, later studies have found them to be indeterminate.[39]
- Teeth from the Alcantara Formation of Brazil were placed in Carcharodontosaurus in 2002,[40] but this has been disputed based on its geographic origin.[10]
Description

Size
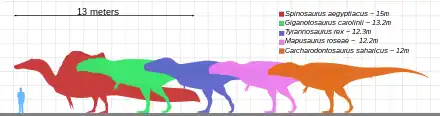
Stromer hypothesized that C. saharicus was around the same size as the tyrannosaurid Gorgosaurus, which would place it at around 8–9 metres (26–30 ft) long, based on his specimen IPHG 1922 X46. This individual was around 15% smaller than the neotype,[41] the latter was estimated to be 12–12.5 metres (39–41 ft) in length and approximately 6–6.2 metric tons (6.6–6.8 short tons) in body mass.[42][27][43] This makes Carcharodontosaurus saharicus one of the largest known theropod dinosaurs and terrestrial carnivores known, being the 3rd biggest carcharodontosaurid and 5th largest theropod overall according to most estimates.[44][27] C. iguidensis was much smaller, only reaching 10 metres (33 ft) in length and 4 metric tons (4.4 short tons) in body mass.[27]
Skull

The largest and most complete skull of C. saharicus would measure 1.6 metres (5.2 ft) when complete, around the same size as the largest Tyrannosaurus skulls. No skulls of the genus preserve premaxillae, complete posterior skull regions, or mandibles. Skulls of carcharodontosaurids tend to be more slender and lightly built than those of later tyrannosaurids, which have robust builds and adaptations for crushing. The neotype cranium tapers towards the front in side view creating a triangular outline. This is similar to that of other carcharodontosaurids like Mapusaurus and Giganotosaurus. Its skull was lighter than that of tyrannosaurids, with the antorbital fenestra composing over 30% of the total skull length as well as being surrounded by fossae in the maxillae (upper jaw bone), nasals (nose bone), jugals (cheekbone), and lacrimals (front orbit bone). Akin to other genera, its nasal is elongated and bears an anterior face covered in a rugose surface. These bumps were likely extended by keratin sheaths, creating a horn-like structure as in Ceratosaurus. A similar rugosity is found on the lacrimal which would also be lengthened by keratin, forming a similar element.[12][7] The most distinctive trait of Carcharodontosaurus' skull is the sculpted exterior of the maxillae , which is unique to the genus. However, C. iguidensis has antorbital fossae limited to the proximity of the antorbital fenestra, a crest running along the medial face of the maxilla, and a process along its midline. These traits are missing in C. saharicus, differentiating the two species.[8]


The maxilla of IPHG 1922 X46 would have been 70 centimetres (28 in) long when complete, whereas the neotype's complete maxilla is much larger. 14 teeth sockets are present in each maxilla. Parts of the braincase are known though much of their morphology is the same as Giganotosaurus'. However, C. saharicus has a much more prominent nuchal crest, which overhangs the skull roof. The frontal bones are firmly fused, a characteristic evident in most theropods.[45] The jugals are broad and triangle-shaped. The lower jaw articulation was placed farther back behind the occipital condyle (where the neck is attached to the skull) compared to other theropods.[7] Two dentary (lower jaw bone) fragments which were referred to C. saharicus by Ibrahim et al (2020) have deep and expanded alveoli (tooth sockets), traits found in other large theropods.[19][12] If like Tyrannotitan and Giganotosaurus, the dentary would have 16 alveoli (tooth sockets).[46]
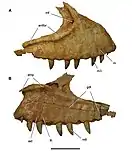



Estimations of the tooth count of Carcharodontosaurus vary, but a recent estimate of 30 dentary, 8 premaxillary, and 24 maxillary teeth for a total of 62 teeth was made.[47] Carcharodontosaurid teeth are some of the largest of any dinosaur group, with a maxillary tooth from IPHG 1922 X46 being 6.8 centimetres (2.7 in) tall and 3.5 centimetres (1.4 in) wide.[13][48] However, they are extremely thin, with most being under a centimeter thick. Serrations are numerous on the anterior and posterior margins, with over 18 to 20 serrations per centimeter of edge in C. saharicus and up to 32 per centimeter in C. iguidensis.[8][13] Its teeth are straight, laterally flattened, and spindle-shaped in cross-section. However, dentition towards the back of the mouth became more recurved than those in the maxilla. The posterior margin of these crowns are recurved and convex at its termination. Bowed enamel wrinkles are present on both dorsoventral sides of the crowns. These wrinkles curve towards the marginal serrations, composing a band-shape along the ends.[7][49][46]
Brain and inner ear
In 2001, Hans C. E. Larsson published a description of the inner ear and endocranium of Carcharodontosaurus saharicus. Starting from the portion of the brain closest to the tip of the animal's snout is the forebrain, which is followed by the midbrain. The midbrain is angled downwards at a 45-degree angle and towards the rear of the animal. This is followed by the hindbrain, which is roughly parallel to the forebrain and forms a roughly 40-degree angle with the midbrain. Overall, the brain of C. saharicus would have been similar to that of a related dinosaur, Allosaurus fragilis. Larsson found that the ratio of the cerebrum to the volume of the brain overall in Carcharodontosaurus was typical for a non-avian reptile. Carcharodontosaurus also had a large optic nerve.[50]
The three semicircular canals of the inner ear of Carcharodontosaurus saharicus—when viewed from the side—had a subtriangular outline. This subtriangular inner-ear configuration is present in Allosaurus, lizards, and turtles, but not in birds.[50] The semi-"circular" canals themselves were very linear, which explains the pointed silhouette. In life, the floccular lobe of the brain would have projected into the area surrounded by the semicircular canals, just like in other non-avian theropods, birds, and pterosaurs.[50]
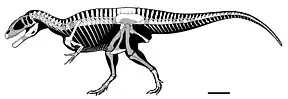
Postcrania
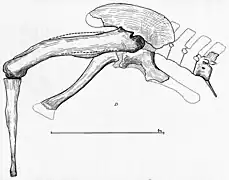
Few postcranial elements are confidently known from Carcharodontosaurus, though many isolated bones from the Sahara have been referred to the genus without detailed study.[8][12][28][9] Like other carcharodontosaurids, it was robust with small forelimbs, an elongated tail, and short neck. The most complete specimen was IPHG 1922 X46, but it was destroyed. This specimen preserved 3 cervical vertebrae, which were weathered severely. One is an axis and the other two are articulating anterior cervicals which are longer and wider than the axis. The cervical vertebrae of Carcharodontosaurus are stout and opisthocoelus (concave posterior ends).[51][46] Cervical vertebrae in this genus, as in Giganotosaurus, are topped by low neural spines joined with sturdy transverse processes which hung over the pleurocoels (shallow depressions on the sides of centra), which would contain pneumatic air sacs to lighten the vertebrae. The centra of these vertebrae are adorned by keels along their ventral sides. An anterior caudal vertebra was also known, which was platycoelous (flat anterior and posterior ends) and short. This caudal was incomplete, missing much of the neural spine, but had diapophyses that would conjugate with the chevrons. The sides of its centrum were pleurocoelus as well. Two blade-like chevrons were preserved in this individual as well.[13]
The pelvis was incomplete, containing both pubes and the left ischium, though complete pelves are known in related genera. The ischium pointed backwards whereas the pubes pointed forwards, a diagnostic trait of saurischians. The pubes were likely nearly 1 metre (3.3 ft) when fully preserved, with shafts that were thin but were transversely expanded at the anterior ends where they connected, creating a V-shape in anterior view. Both femora in addition to the left fibula were recovered, the former element being one of the largest recorded from a theropod at 1.26 metres (4.1 ft) in length. Its femora lacked strong curvature and are mostly straight except for the anterior and posterior ends. The greater trochanter is small but has a notable protrusion, which would attach to the m. caudofemoralis longus muscle of the tail. Its fibula was only 88 centimetres (35 in) long, around 1/3rd the length of the femora. The anterior end was triangular in lateral view with bulging condyles whereas the posterior end is rounded.[13]
Classification
Carcharodontosaurus is the type genus of the family Carcharodontosauridae and subfamily Carcharodontosaurinae. This subfamily contains Carcharodontosaurus itself as well as the other carcharodontosaurines Giganotosaurus, Mapusaurus, Meraxes, and Tyrannotitan; however, these genera make up an independent tribe: Giganotosaurini. Carcharodontosauridae was a clade created by Stromer for Carcharodontosaurus and Bahariasaurus, though the name remained unused until the recognition of other members of the group in the late 20th century. He noted the likeness of Carcharodontosaurus bones to the American theropods Allosaurus and Tyrannosaurus, leading him to consider the family part of Theropoda.[14][13]
Paul Sereno's description of Carcharodontosaurus fossils in 1996 led to the realization of a transcontinental clade of carcharodontosaurids. As more carcharodontosaurids were discovered, their interrelationships became even clearer. The group was defined as all allosauroids closer to Carcharodontosaurus than Allosaurus or Sinraptor by the paleontologist Thomas R. Holtz and colleagues in 2004.[52] Carcharodontosaurus is more poorly known than most other carcharodontosaurids, with Meraxes and Giganotosaurus represented by nearly complete skeletons.[53][54] Members of the family have been recognized from the Late Jurassic to Middle Cretaceous of every continent except Oceania and Antarctica.[44][55][7]
Canale et al. (2022) recovered Carcharodontosaurus as the earliest diverging member of Carcharodontosaurinae. The cladogram results of their phylogenetic analyses are displayed in the cladogram below:[53]
| Carcharodontosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Evolution
Coria and Salgado suggested that the convergent evolution of gigantism in theropods could have been linked to common conditions in their environments or ecosystems.[54] Sereno and colleagues found that the presence of carcharodontosaurids in Africa (Carcharodontosaurus), North America (Acrocanthosaurus), and South America (Giganotosaurus), showed the group had a transcontinental distribution by the Early Cretaceous period. Dispersal routes between the northern and southern continents appear to have been severed by ocean barriers in the Late Cretaceous, which led to more distinct, provincial faunas, by preventing exchange.[56][7] Previously, it was thought that the Cretaceous world was biogeographically separated, with the northern continents being dominated by tyrannosaurids, South America by abelisaurids, and Africa by carcharodontosaurids.[57] The subfamily Carcharodontosaurinae, in which Carcharodontosaurus belongs, appears to have been restricted to the southern continent of Gondwana (formed by South America and Africa), where they were probably the apex predators.[52] The South American tribe Giganotosaurini may have been separated from their African relatives through vicariance, when Gondwana broke up during the Aptian–Albian ages of the Early Cretaceous.[58]
Paleobiology
Lifting capabilities
A biomechanical analysis of Carcharodontosaurus' lifting capabilities was conducted by paleontologist Donald Henderson and paleoartist Robert Nicholls in 2015. The authors used 3D models of the animal as well as a subadult sauropod Limaysaurus, which although not found alongside Carcharodontosaurus, is similar to the rebbachisaurids of the Kem Kem Beds. The models included the size of the lungs and other pneumatic structures of the two, fostering an accurate weight simulation of the scenario. Henderson & Nicholls' study found that an adult C. saharicus could hold a maximum of 424 kg (935 lb), half the weight of an adult Limaysaurus. However, two C. saharicus adults could together lift as much as 850 kilograms (1,870 lb).[42]
Feeding and diet

The dentition of allosauroids is distinct, with carcharodontosaurid teeth bearing distinctly thin and blade-like teeth. However, these teeth are thin and likely could not sustain impact against hard surfaces like bone without potentially bending and snapping. This danger is exacerbated by the straight edges, slightly recurved tips, and sinusoidal shapes observed in their dentition. Despite these traits, the teeth are still much more robust than those of smaller theropods and due to their overall size could take more pressure. Carcharodontosaurus also had a high tooth replacement rate meaning that damaged teeth could be replaced easily in contrast to extant bone-crushing mammals who spend much of their energy maintaining their teeth.[59][60] Evidence of bone-crunching bites is observed in Allosaurus, which would engage in ritual face-biting with other individuals and bite into the pelves of Stegosaurus as shown by bite marks.[61][62][63]
Bite forces of Carcharodontosaurus as well as other giant theropods including Acrocanthosaurus and Tyrannosaurus have been analyzed. Studies reported that carcharodontosaurids had much lower bite forces than Tyrannosaurus despite being in the same size class. The anterior bite force of C. saharicus was estimated in a 2022 paper to be 11,312 newtons while the posterior bite force was 25,449 newtons. This is much lower than that of Tyrannosaurus, implying that it did not eat bones.[64][65] Finite element accounts of the skulls of theropods have also been taken, which further supported the idea that Carcharodontosaurus ate softer food than tyrannosaurids. Great amounts of stress were recovered in the posterior part of the cranium near the quadrate in Carcharodontosaurus, Spinosaurus, and Acrocanthosaurus. The skulls of these theropods had higher relative stress quantities in opposition to that of smaller genera. This indicates that the crania of giant taxa (ex. Carcharodontosaurus) were unstable due to having large pneumatic structures to save weight instead of creating a firm build. However, Spinosaurus and Suchomimus experienced even greater values of stress meaning that they could only consume light, small prey instead of larger items, which the stronger skull of Carcharodontosaurus could bite while sustaining the stress.[66]
_3_(15375691822).jpg.webp)
_2_(15352983706).jpg.webp)
Isotopic analyses of the teeth of C. saharicus have found δ18O values that are higher than that of the contemporary Spinosaurus, suggesting the latter pursued semi-aquatic habits whereas Carcharodontosaurus was more terrestrial.[67] This is further supported by the taphonomy of C. saharicus teeth, which are more often found in land terrains than aquatic ones.[47] Carcharodontosaurus was also a homeotherm with an endotherm-like thermophysiology as inferred by these isotopes meaning that most of its oxygen was accumulated by drinking water rather than being in it.[68][69]
Crest function
Theropods such as Carcharodontosaurus, Allosaurus, and Acrocanthosaurus have enlarged lacrimal crests, whose purpose is unknown. Paleontologist Daniel Chure hypothesized that these crests were used for "head-butting" between individuals, but how durable they are has not been studied.[70]
Vision
A 2006 study by biologist Kent Stevens analyzed the binocular vision capabilities of the allosauroids Carcharodontosaurus and Allosaurus as well as several coelurosaurs including Tyrannosaurus and Stenonychosaurus. By applying modified perimetry to models of these dinosaurs' heads, Stevens deduced that the binocular vision of Carcharodontosaurus was limited, a side effect of its large, elongated rostrum. Its greatest degree of binocular vision was at higher elevations, suggesting that Carcharodontosaurus may have habitually held its head at a downward 40° angle with its eyes facing up accordingly to achieve maximum binocular vision. The range of vision seen in these allosauroids is comparable to that of crocodiles, suggesting that they were ambush predators. They likely sensed prey via motion parallax between prey and background, with a narrow binocular field of vision helping predators judge prey distances and time attacks.[71]
Pathology
The neotype skull of C. saharicus is one of many allosauroid individuals to preserve pathologies, with signs of biting, infection, and breaks observed in Allosaurus and Acrocanthosaurus among others.[72][73] This skull bears a circular puncture wound in the nasal and "an abnormal projection of bone on the antorbital rim".[73] A later study theorized that this was the result of craniofacial bites.[74]
Paleoenvironment

Fossils of Carcharodontosaurus are known from several Cretaceous-age sites across North Africa, similar to the ranges of Spinosaurus and Deltadromeus.[19][7] North Africa during this period bordered the Tethys Sea, which transformed the region into a mangrove-dominated coastal environment filled with vast tidal flats and waterways.[75][76][12] Isotopes from Carcharodontosaurus and Spinosaurus fossils suggest that the Kem Kem Beds witnessed a temporary monsoon season rather than constant rainfall, similar to modern conditions present in sub-tropical and tropical environments in Southeast Asia and Sub-Saharan Africa.[67][77] These riverine deposits bore large fishes, including the sawskate Onchopristis, coelacanth Mawsonia, and bichir Bawitius.[12] This led to an abundance of piscivorous crocodyliformes evolving in response, such as the giant stomatosuchid Stomatosuchus in Egypt and the genera Elosuchus, Laganosuchus, and Aegisuchus from Morocco.[78][12] Morocco also bore an abundance of pterosaurs like Siroccopteryx and Nicorhynchus.[79][80]
The composition of the dinosaur fauna of these sites is an anomaly, as there are fewer herbivorous dinosaur species relative to carnivorous dinosaurs than usual. This indicates that there was niche partitioning between the different theropod clades, with spinosaurids consuming fish while other groups hunted herbivorous dinosaurs.[81] Isotopic evidence supports this, which found greater quantities of sizable, terrestrial animals in the diets of carcharodontosaurids and ceratosaurs from both the Kem Kem Beds and Elrhaz Formation.[82][9] Some sauropods are known from the Bahariya Formation such as Paralititan and Aegyptosaurus,[83] while Rebbachisaurus is found in the Kem Kem Beds.[84] Carcharodontosaurids are represented by C. saharicus and Sauroniops in the Kem Kem Beds, Eocarcharia and potentially Carcharodontosaurus in the Elrhaz Formation, and C. iguidensis in the Echkar Formation.[8]
References
- Benyoucef, Madani; Pérez-García, Adán; Bendella, Mohamed; Ortega, Francisco; Vullo, Romain; Bouchemla, Imad; Ferré, Bruno (2022). "The "mid"-Cretaceous (Lower Cenomanian) Continental Vertebrates of Gara Samani, Algeria. Sedimentological Framework and Palaeodiversity". Frontiers in Earth Science. 10: 927059. Bibcode:2022FrEaS..10.7059B. doi:10.3389/feart.2022.927059. ISSN 2296-6463.
- Depéret, Charles; Savornin, Justin (1927). "La faune de reptiles et de poisons albiens de Timimoun (Sahara algérien)". Bulletin de la Société Géologique de France. 27: 257–265.
- Buffetaut, E., & Ouaja, M. (2002). A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae. Bulletin de la Société Géologique de France, 173(5), 415-421.
- Depéret, Charles; Savornin, Justin (1925). "Sur la découverte d'une faune de vertébrés albiens à Timimoun (Sahara occidental)". Comptes Rendus de l'Académie des Sciences. 181: 1108–1111.
- Benson, Roger B. J.; Barrett, Paul M.; Powell, H. Philip; Norman, David B. (2008). "The taxonomic status of Megalosaurus bucklandii (Dinosauria, Theropoda) from the Middle Jurassic of Oxfordshire, UK". Palaeontology. 51 (2): 419–424. Bibcode:2008Palgy..51..419B. doi:10.1111/j.1475-4983.2008.00751.x. S2CID 83324840.
- Huene, Friedrich (1956). "Palaeontologie und Phylogenie der Niederen Tetrapoden". VEB Gustav Fischer Verlang, Jena.: 1–716.
- Sereno, Paul C.; Dutheil, Didier B.; Iarochene, M.; Larsson, Hans C. E.; Lyon, Gabrielle H.; Magwene, Paul M.; Sidor, Christian A.; Varricchio, David J.; Wilson, Jeffrey A. (May 17, 1996). "Predatory Dinosaurs from the Sahara and Late Cretaceous Faunal Differentiation". Science. 272 (5264): 986–991. Bibcode:1996Sci...272..986S. doi:10.1126/science.272.5264.986. ISSN 0036-8075. PMID 8662584. S2CID 39658297.
- Brusatte, Stephen L.; Sereno, Paul C. (December 12, 2007). "A new species of Carcharodontosaurus (Dinosauria: Theropoda) from the Cenomanian of Niger and a revision of the genus". Journal of Vertebrate Paleontology. 27 (4): 902–916. doi:10.1671/0272-4634(2007)27[902:ANSOCD]2.0.CO;2. ISSN 0272-4634. S2CID 86202969.
- de Lapparent, Albert (1960). "Les dinosauriens du "Continental intercalaire" du Sahara central" (PDF). Memoirs of the Geological Society of France. 88A: 1–57.
- Mortimer, Mickey (2023). "Carnosauria". The Theropod Database. Archived from the original on May 28, 2023. Retrieved June 6, 2023.
- Gabani, A.; Mammeri, C.; Adaci, M.; Bensalah, M.; Mahboubi, M. (2016). "Le Crétacé continental à vertébrés de la bordure sud du Plateau de Tinhert: considérations stratigraphiques et bilan paléontologique". Mémoire du Service Géologique de l'Algérie. 19: 39–61.
- Ibrahim, Nizar; Sereno, Paul C.; Varricchio, David J.; Martill, David M.; Dutheil, Didier B.; Unwin, David M.; Baidder, Lahssen; Larsson, Hans C. E.; Zouhri, Samir; Kaoukaya, Abdelhadi (April 21, 2020). "Geology and paleontology of the Upper Cretaceous Kem Kem Group of eastern Morocco". ZooKeys (928): 1–216. doi:10.3897/zookeys.928.47517. ISSN 1313-2970. PMC 7188693. PMID 32362741.
- Stromer, Ernst (1931). "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharîjestufe (unterstes Cenoman). 10. Ein Skelett-Rest von Carcharodontosaurus nov. gen". Abhandlungen der Bayerischen Akademie der Wissenschaften Mathematisch-naturwissenschaftliche Abteilung. Neue Folge (in German). 9: 1–23.
- Stromer, E. (1934). "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 13. Dinosauria". Abhandlungen der Bayerischen Akademie der Wissenschaften Mathematisch-naturwissenschaftliche Abteilung. Neue Folge (in German). 22: 1–79.
- "Rocky Road: Ernst Stromer". www.strangescience.net. Retrieved July 8, 2023.
- Smith, Joshua B.; Lamanna, Matthew C.; Mayr, Helmut; Lacovara, Kenneth J. (2006). "New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915". Journal of Paleontology. 80 (2): 400–406. doi:10.1666/0022-3360(2006)080[0400:NIRTHO]2.0.CO;2. ISSN 0022-3360. S2CID 130989487.
- Nothdurft, William; Smith, Josh (September 24, 2002). The Lost Dinosaurs of Egypt. Random House Publishing Group. ISBN 978-1-58836-117-2.
- Chiarenza, Alfio Alessandro; Cau, Andrea (February 29, 2016). "A large abelisaurid (Dinosauria, Theropoda) from Morocco and comments on the Cenomanian theropods from North Africa". PeerJ. 4: e1754. doi:10.7717/peerj.1754. ISSN 2167-8359. PMC 4782726. PMID 26966675.
- Russell, Dale (1996). "Isolated Dinosaur bones from the Middle Cretaceous of the Tafilalt, Morocco". Bulletin du Muséum national d'Histoire naturelle, 4ème série – section C – Sciences de la Terre, Paléontologie, Géologie, Minéralogie (in French). 18 (2–3).
- Ibrahim, N., Sereno, P. C., Zouhri, S., & Zouhri, S. (2017). Les dinosaures du Maroc–aperçu historique et travaux récents. Mémoires de la Societé géologique de France, 180, 249-284.
- Lavocat, Rene (1954). "Sur les dinosauriens du Continental Intercalaire des Kem-Kem de la Daoura". Comptes Rendus 19th International Geological Congress. 1952: 65–68.
- Sereno, Paul C.; Myhrvold, Nathan; Henderson, Donald M.; Fish, Frank E.; Vidal, Daniel; Baumgart, Stephanie L.; Keillor, Tyler M.; Formoso, Kiersten K.; Conroy, Lauren L. (November 30, 2022). Zhu, Min; Rutz, Christian; Zhu, Min; Holtz, Thomas R.; Hone, David (eds.). "Spinosaurus is not an aquatic dinosaur". eLife. 11: e80092. doi:10.7554/eLife.80092. ISSN 2050-084X. PMC 9711522. PMID 36448670.
- Cau, Andrea; Dalla Vecchia, Fabio M.; Fabbri, Matteo (March 1, 2013). "A thick-skulled theropod (Dinosauria, Saurischia) from the Upper Cretaceous of Morocco with implications for carcharodontosaurid cranial evolution". Cretaceous Research. 40: 251–260. Bibcode:2013CrRes..40..251C. doi:10.1016/j.cretres.2012.09.002. ISSN 0195-6671.
- Candeiro, Carlos Roberto dos Anjos; Brusatte, Stephen Louis; Vidal, Luciano; Pereira, Paulo Victor Luiz Gomes da Costa (July 26, 2018). "Paleobiogeographic evolution and distribution of Carcharodontosauridae (Dinosauria, Theropoda) during the middle Cretaceous of North Africa". Papéis Avulsos de Zoologia. 58: e20185829. doi:10.11606/1807-0205/2018.58.29. ISSN 0031-1049. S2CID 53353652.
- Paterna, Alessandro; Cau, Andrea (October 11, 2022). "New giant theropod material from the Kem Kem Compound Assemblage (Morocco) with implications on the diversity of the mid-Cretaceous carcharodontosaurids from North Africa". Historical Biology: 1–9. doi:10.1080/08912963.2022.2131406. ISSN 0891-2963. S2CID 252856791.
- Figueiredo (1998). "Os dinossáurios carnívoros: A sua descrição e modo de vida". Centro Portugues de Geo-historia e Prehistoria. pp. 1–4.
- Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press. pp. 103–104. ISBN 978-1-78684-190-2. OCLC 985402380.
- Taquet, Philippe (1976). Géologie et Paléontologie du Gisement de Gadoufaoua (Aptien du Niger). Paris: Cahiers de Paléontologie, Centre National de la Recherche Scientifique, Paris. p. 53.
- Ibrahim, Nizar; Sereno, Paul C.; Dal Sasso, Cristiano; Maganuco, Simone; Fabbri, Matteo; Martill, David M.; Zouhri, Samir; Myhrvold, Nathan; Iurino, Dawid A. (September 26, 2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–1616. Bibcode:2014Sci...345.1613I. doi:10.1126/science.1258750. ISSN 0036-8075. PMID 25213375. S2CID 34421257.
- Schluter; Schwarzhans (1978). "Eine Bonebed-Lagerstatte aus dem Wealden Sud-Tunesiens (Umgebung Ksar Krerachfa)". Berliner geowissenchaftliche Abhandlungen A. 8: 53–65.
- Fanti, Federico; Cau, Andrea; Martinelli, Agnese; Contessi, Michela (September 15, 2014). "Integrating palaeoecology and morphology in theropod diversity estimation: A case from the Aptian-Albian of Tunisia". Palaeogeography, Palaeoclimatology, Palaeoecology. 410: 39–57. Bibcode:2014PPP...410...39F. doi:10.1016/j.palaeo.2014.05.033. ISSN 0031-0182.
- Churcher, C. S. (1999). A note on the Late Cretaceous vertebrate fauna of the Dakhleh Oasis. OXBOW MONOGRAPH, 55-68.
- Singer (2015). "JuraPark na tropie nowych dinozaurow z Maroka". Jurapark. Archived from the original on December 6, 2015. Retrieved June 27, 2023.
- Molina-Pérez, Rubén; Larramendi, Asier; Connolly, David; Cruz, Gonzalo Ángel Ramírez (June 25, 2019). Dinosaur Facts and Figures: The Theropods and Other Dinosauriformes. Princeton University Press. ISBN 978-0-691-19059-4.
- Crusafont-Pairó, M., & Adrover, R. (1966). El primer representante de la clase mamíferos hallado en el Mesozoico de España. Teruel, 35, 139-143.
- Kuhne, W. G.; Crusafont-Pairo, M. (1968). "Mamíferos del Wealdiense de Uña, cerca de Cuenca". Acta Geológica Hispánica. 3 (5): 133–134.
- Gascó, Francisco; Cobos, Alberto; Royo-Torres, Rafael; Mampel, Luis; Alcalá, Luis (June 1, 2012). "Theropod teeth diversity from the Villar del Arzobispo Formation (Tithonian–Berriasian) at Riodeva (Teruel, Spain)". Palaeobiodiversity and Palaeoenvironments. 92 (2): 273–285. doi:10.1007/s12549-012-0079-3. ISSN 1867-1608. S2CID 129930988.
- Bond, Geoffrey; Bromley, K. (December 1, 1970). "Sediments with the remains of dinosaurs near Gokwe, Rhodesia". Palaeogeography, Palaeoclimatology, Palaeoecology. 8 (4): 313–327. Bibcode:1970PPP.....8..313B. doi:10.1016/0031-0182(70)90104-5. ISSN 0031-0182.
- Munyikwa, D.; Sampson, S. D.; Rogers, R. R.; Forster, C. A.; Curry, K. A.; Curtice, B. D. (1998). "Vertebrate palaeontology and geology of the Gokwe Formation, Zimbabwe". Journal of African Earth Sciences. 27 (1): 142.
- Medeiros, Manuel; Schultz, Cesar (2002). "A fauna dinossauriana da "Laje do Coringa", Cretaceo medio de Nordeste do Brasil". Arquivos do Museo Nacional, Rio de Janeiro. 60 (3): 155–162.
- Nothdurft & Smith 2002, p. 109.
- Henderson, D.M.; Nicholls, R. (2015). "Balance and Strength—Estimating the Maximum Prey-Lifting Potential of the Large Predatory Dinosaur Carcharodontosaurus saharicus". The Anatomical Record. 298 (8): 1367–1375. doi:10.1002/ar.23164. PMID 25884664. S2CID 19465614.
- Seebacher, F. (2001). "A New Method to Calculate Allometric Length-Mass Relationships of Dinosaurs" (PDF). Journal of Vertebrate Paleontology. 21 (1): 51–60. CiteSeerX 10.1.1.462.255. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2. ISSN 0272-4634. S2CID 53446536.
- Coria, Rodolfo A.; Currie, Philip J.; Ortega, Francisco; Baiano, Mattia A. (July 1, 2020). "An Early Cretaceous, medium-sized carcharodontosaurid theropod (Dinosauria, Saurischia) from the Mulichinco Formation (upper Valanginian), Neuquén Province, Patagonia, Argentina". Cretaceous Research. 111: 104319. Bibcode:2020CrRes.11104319C. doi:10.1016/j.cretres.2019.104319. hdl:11336/122794. ISSN 0195-6671. S2CID 214475057.
- Coria, Rodolfo A.; Currie, Philip J. (January 14, 2003). "The braincase of Giganotosaurus carolinii (Dinosauria: Theropoda) from the Upper Cretaceous of Argentina". Journal of Vertebrate Paleontology. 22 (4): 802–811. doi:10.1671/0272-4634(2002)022[0802:TBOGCD]2.0.CO;2. ISSN 0272-4634. S2CID 85678725.
- Novas, Fernando E.; de Valais, Silvina; Vickers-Rich, Pat; Rich, Tom (May 1, 2005). "A large Cretaceous theropod from Patagonia, Argentina, and the evolution of carcharodontosaurids". Naturwissenschaften. 92 (5): 226–230. Bibcode:2005NW.....92..226N. doi:10.1007/s00114-005-0623-3. ISSN 1432-1904. PMID 15834691. S2CID 24015414.
- Beevor, Thomas; Quigley, Aaron; Smith, Roy E.; Smyth, Robert S. H.; Ibrahim, Nizar; Zouhri, Samir; Martill, David M. (January 1, 2021). "Taphonomic evidence supports an aquatic lifestyle for Spinosaurus". Cretaceous Research. 117: 104627. Bibcode:2021CrRes.11704627B. doi:10.1016/j.cretres.2020.104627. ISSN 0195-6671. S2CID 224888268.
- "Discoveries | Paul Sereno - Paleontologist | The University of Chicago". paulsereno.uchicago.edu. Retrieved June 30, 2023.
- Brusatte, Stephen L.; Benson, Roger B. J.; Carr, Thomas D.; Williamson, Thomas E.; Sereno, Paul C. (December 12, 2007). "The systematic utility of theropod enamel wrinkles". Journal of Vertebrate Paleontology. 27 (4): 1052–1056. doi:10.1671/0272-4634(2007)27[1052:TSUOTE]2.0.CO;2. ISSN 0272-4634. S2CID 85615205.
- Larsson, H.C.E. 2001. Endocranial anatomy of Carcharodontosaurus saharicus (Theropoda: Allosauroidea) and its implications for theropod brain evolution. pp. 19–33. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- Harris, Jerald D. (1998). "A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiological implications, based on a new specimen from Texas". New Mexico Museum of Natural History and Science Bulletin. 13: 1–75.
- Novas, Fernando E.; Agnolín, Federico L.; Ezcurra, Martín D.; Porfiri, Juan; Canale, Juan I. (October 1, 2013). "Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia". Cretaceous Research. 45: 174–215. Bibcode:2013CrRes..45..174N. doi:10.1016/j.cretres.2013.04.001. ISSN 0195-6671.
- Canale, Juan I.; Apesteguía, Sebastián; Gallina, Pablo A.; Mitchell, Jonathan; Smith, Nathan D.; Cullen, Thomas M.; Shinya, Akiko; Haluza, Alejandro; Gianechini, Federico A.; Makovicky, Peter J. (July 7, 2022). "New giant carnivorous dinosaur reveals convergent evolutionary trends in theropod arm reduction". Current Biology. 32 (14): 3195–3202.e5. doi:10.1016/j.cub.2022.05.057. ISSN 0960-9822. PMID 35803271. S2CID 250343124.
- Coria, Rodolfo A.; Salgado, Leonardo (1995). "A new giant carnivorous dinosaur from the Cretaceous of Patagonia". Nature. 377 (6546): 224–226. Bibcode:1995Natur.377..224C. doi:10.1038/377224a0. ISSN 1476-4687. S2CID 30701725.
- Brusatte, Stephen L.; Benson, Roger B.J.; Xu, Xing (2012). "A Reassessment ofKelmayisaurus petrolicus, a Large Theropod Dinosaur from the Early Cretaceous of China". Acta Palaeontologica Polonica. 57 (1): 65–72. doi:10.4202/app.2010.0125. ISSN 0567-7920. S2CID 53387460.
- Currie, Philip J. (May 17, 1996). "Out of Africa: Meat-Eating Dinosaurs That Challenge Tyrannosaurus rex". Science. 272 (5264): 971–972. Bibcode:1996Sci...272..971C. doi:10.1126/science.272.5264.971. ISSN 0036-8075. S2CID 85110425.
- Coria, Rodolfo A.; Salgado, Leonardo (June 1996). "Dinosaurios carnívoros de Sudamérica". Investigación y Ciencia (in Spanish) (237): 39–40.
- Canale, J. I.; Novas, F. E.; Pol, D. (2014). "Osteology and phylogenetic relationships of Tyrannotitan chubutensis Novas, de Valais, Vickers-Rich and Rich, 2005 (Theropoda: Carcharodontosauridae) from the Lower Cretaceous of Patagonia, Argentina". Historical Biology. 27 (1): 1–32. doi:10.1080/08912963.2013.861830. hdl:11336/17607. S2CID 84583928.
- Van Valkenburgh, Blaire (1988). "Incidence of Tooth Breakage Among Large, Predatory Mammals". The American Naturalist. 131 (2): 291–302. doi:10.1086/284790. ISSN 0003-0147. S2CID 222330098.
- Van Valkenburgh, Blaire (December 17, 2008). "Costs of carnivory: tooth fracture in Pleistocene and Recent carnivorans: COSTS OF CARNIVORY". Biological Journal of the Linnean Society. 96 (1): 68–81. doi:10.1111/j.1095-8312.2008.01108.x. S2CID 85623961.
- Hone, David W. E.; Rauhut, Oliver W. M. (2010). "Feeding behaviour and bone utilization by theropod dinosaurs". Lethaia. 43 (2): 232–244. doi:10.1111/j.1502-3931.2009.00187.x. ISSN 0024-1164.
- Tanke, D.; Currie, P. J. (1998). "Head-biting behavior in theropod dinosaurs: Paleopathological evidence". ERA. doi:10.7939/R34T6FJ1P. S2CID 90552600. Retrieved July 2, 2023.
- Drumheller, Stephanie K.; McHugh, Julia B.; Kane, Miriam; Riedel, Anja; D’Amore, Domenic C. (May 27, 2020). "High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem". PLOS ONE. 15 (5): e0233115. Bibcode:2020PLoSO..1533115D. doi:10.1371/journal.pone.0233115. ISSN 1932-6203. PMC 7252595. PMID 32459808.
- Sakamoto, Manabu (July 12, 2022). "Estimating bite force in extinct dinosaurs using phylogenetically predicted physiological cross-sectional areas of jaw adductor muscles". PeerJ. 10: e13731. doi:10.7717/peerj.13731. ISSN 2167-8359. PMC 9285543. PMID 35846881.
- Gignac, Paul M.; Erickson, Gregory M. (May 17, 2017). "The Biomechanics Behind Extreme Osteophagy in Tyrannosaurus rex". Scientific Reports. 7 (1): 2012. Bibcode:2017NatSR...7.2012G. doi:10.1038/s41598-017-02161-w. ISSN 2045-2322. PMC 5435714. PMID 28515439.
- Rayfield, Emily (2011). "Structural performance of tetanuran theropod skulls, with emphasis on the Megalosauridae, Spinosauridae and Carcharodontosauridae" (PDF). Special Papers in Palaeontology. 86: 241–253.
- Goedert, J; Amiot, R; Boudad, L; Buffetaut, E; Fourel, F; Godefroit, P; Kusuhashi, N; Suteethorn, V; Tong, H; Watabe, M; Lecuyer, C (2016). "Preliminary investigation of seasonal patterns recorded in the oxygen isotope compositions of theropod dinosaur tooth enamel". PALAIOS. 31 (1): 10–19. Bibcode:2016Palai..31...10G. doi:10.2110/palo.2015.018. S2CID 130878403.
- Amiot, Romain; Wang, Xu; Lécuyer, Christophe; Buffetaut, Eric; Boudad, Larbi; Cavin, Lionel; Ding, Zhongli; Fluteau, Frédéric; Kellner, Alexander W.A.; Tong, Haiyan; Zhang, Fusong (2010). "Oxygen and carbon isotope compositions of middle Cretaceous vertebrates from North Africa and Brazil: Ecological and environmental significance". Palaeogeography, Palaeoclimatology, Palaeoecology. 297 (2): 439–451. Bibcode:2010PPP...297..439A. doi:10.1016/j.palaeo.2010.08.027.
- Amiot, Romain; Lécuyer, Christophe; Buffetaut, Eric; Escarguel, Gilles; Fluteau, Frédéric; Martineau, François (June 15, 2006). "Oxygen isotopes from biogenic apatites suggest widespread endothermy in Cretaceous dinosaurs". Earth and Planetary Science Letters. 246 (1): 41–54. Bibcode:2006E&PSL.246...41A. doi:10.1016/j.epsl.2006.04.018. ISSN 0012-821X.
- Chure, Daniel (2000). "On the orbit of theropod dinosaurs". Gaia. 15: 233–240.
- Stevens, Kent A. (June 12, 2006). "Binocular vision in theropod dinosaurs". Journal of Vertebrate Paleontology. 26 (2): 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2. ISSN 0272-4634. S2CID 85694979.
- Chinsamy, Anusuya; Tumarkin-Deratzian, Allison (2009). "Pathologic Bone Tissues in a Turkey Vulture and a Nonavian Dinosaur: Implications for Interpreting Endosteal Bone and Radial Fibrolamellar Bone in Fossil Dinosaurs". The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. 292 (9): 1478–1484. doi:10.1002/ar.20991. PMID 19711479. S2CID 41596233.
- Molnar, R. E. (2001). "Theropod paleopathology: a literature survey". Mesozoic vertebrate life: 342.
- Rothschild, Bruce; Tanke, Darren (2005). "Theropod paleopathology: state-of-the-art review". The Carnivorous Dinosaurs (1st ed.). Indianapolis: Indiana University Press. p. 351. ISBN 9780253345394.
- Wanas, Hamdalla A.; Assal, Ehab M. (March 1, 2021). "Provenance, tectonic setting and source area-paleoweathering of sandstones of the Bahariya Formation in the Bahariya Oasis, Egypt: An implication to paleoclimate and paleogeography of the southern Neo-Tethys region during Early Cenomanian". Sedimentary Geology. 413: 105822. Bibcode:2021SedG..41305822W. doi:10.1016/j.sedgeo.2020.105822. ISSN 0037-0738. S2CID 230598476.
- Hamed, Younes; Al-Gamal, Samir Anwar; Ali, Wassim; Nahid, Abederazzak; Dhia, Hamed Ben (March 1, 2014). "Palaeoenvironments of the Continental Intercalaire fossil from the Late Cretaceous (Barremian-Albian) in North Africa: a case study of southern Tunisia". Arabian Journal of Geosciences. 7 (3): 1165–1177. doi:10.1007/s12517-012-0804-2. ISSN 1866-7538. S2CID 128755145.
- Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- Holliday, Casey M.; Gardner, Nicholas M. (January 31, 2012). "A New Eusuchian Crocodyliform with Novel Cranial Integument and Its Significance for the Origin and Evolution of Crocodylia". PLOS ONE. 7 (1): e30471. Bibcode:2012PLoSO...730471H. doi:10.1371/journal.pone.0030471. ISSN 1932-6203. PMC 3269432. PMID 22303441.
- Ibrahim, Nizar; Unwin, David M.; Martill, David M.; Baidder, Lahssen; Zouhri, Samir (May 26, 2010). "A New Pterosaur (Pterodactyloidea: Azhdarchidae) from the Upper Cretaceous of Morocco". PLOS ONE. 5 (5): e10875. Bibcode:2010PLoSO...510875I. doi:10.1371/journal.pone.0010875. ISSN 1932-6203. PMC 2877115. PMID 20520782.
- Borja Holgado; Rodrigo V. Pêgas (2020). "A taxonomic and phylogenetic review of the anhanguerid pterosaur group Coloborhynchinae and the new clade Tropeognathinae". Acta Palaeontologica Polonica. 65 (4): 743–761. doi:10.4202/app.00751.2020. S2CID 222075296.
- Ibrahim, N; Dal Sasso, C; Maganuco, S; Fabbri, M; Martill, D; Gorscak, E; Lamanna, M (2016). "Evidence of a derived titanosaurian (Dinosauria, Sauropoda) in the "Kem Kem beds" of Morocco, with comments on sauropod paleoecology in the Cretaceous of Africa". Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin. 71: 149–159.
- Hassler, A.; Martin, J. E.; Amiot, R.; Tacail, T.; Godet, F. Arnaud; Allain, R.; Balter, V. (April 11, 2018). "Calcium isotopes offer clues on resource partitioning among Cretaceous predatory dinosaurs". Proceedings of the Royal Society B: Biological Sciences. 285 (1876): 20180197. doi:10.1098/rspb.2018.0197. ISSN 0962-8452. PMC 5904318. PMID 29643213.
- Smith, Joshua B.; Lamanna, Matthew C.; Lacovara, Kenneth J.; Dodson, Peter; Smith, Jennifer R.; Poole, Jason C.; Giegengack, Robert; Attia, Yousry (2001). "A Giant Sauropod Dinosaur from an Upper Cretaceous Mangrove Deposit in Egypt". Science. 292 (5522): 1704–1706. Bibcode:2001Sci...292.1704S. doi:10.1126/science.1060561. ISSN 0036-8075. PMID 11387472. S2CID 33454060.
- Wilson, Jeffrey A.; Allain, Ronan (July 4, 2015). "Osteology of Rebbachisaurus garasbae Lavocat, 1954, a diplodocoid (Dinosauria, Sauropoda) from the early Late Cretaceous–aged Kem Kem beds of southeastern Morocco". Journal of Vertebrate Paleontology. 35 (4): e1000701. Bibcode:2015JVPal..35E0701W. doi:10.1080/02724634.2014.1000701. ISSN 0272-4634. S2CID 129846042.
Bibliography
- Nothdurft, W., & Smith, J. (2002). The lost dinosaurs of Egypt. Random House.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||

.jpg.webp)