Cercozoa
Cercozoa (now synonymised with Filosa)[2] is a phylum of diverse single-celled eukaryotes.[4][5] They lack shared morphological characteristics at the microscopic level,[6] and are instead united by molecular phylogenies of rRNA and actin or polyubiquitin.[7] They were the first major eukaryotic group to be recognized mainly through molecular phylogenies.[8] They are the natural predators of many species of bacteria. They are closely related to the phylum Retaria, comprising amoeboids that usually have complex shells, and together form a supergroup called Rhizaria.[2]
| Cercozoa | |
|---|---|
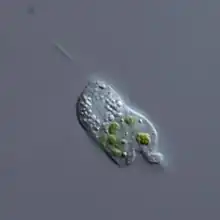 | |
| Cercomonas | |
| Scientific classification | |
| Domain: | Eukaryota |
| Clade: | Diaphoretickes |
| Clade: | SAR |
| Clade: | Rhizaria |
| Phylum: | Cercozoa Cavalier-Smith, 1998[1] emend. Adl et al., 2005 emend. Cavalier-Smith, 2018[2] |
| Classes | |
| |
| Synonyms | |
Characteristics
The group includes most amoeboids and flagellates that feed by means of filose pseudopods. These may be restricted to part of the cell surface, but there is never a true cytostome or mouth as found in many other protozoa. They show a variety of forms[9] and have proven difficult to define in terms of structural characteristics, although their unity is strongly supported by phylogenetic studies.
Diversity
Some cercozoans are grouped by whether they are "filose" or "reticulose" in the behavior of their cytoskeleton when moving:[10]
- Filose, meaning their pseudopods develop as filopodia. For example:
- Euglyphids, filose amoebae with shells of siliceous scales or plates, which are commonly found in soils, nutrient-rich waters, and on aquatic plants.
- Gromia, a shelled amoeba.
- Tectofilosids, filose amoebae that produce organic shells.
- Cercomonads, common soil-dwelling amoeboflagellates.
- Reticulose, meaning they form a reticulating net of pseudopods. For example:
- Chlorarachniophytes, set apart by the presence of chloroplasts bound by four membranes and still possess a vestigial nucleus, called a nucleomorph. As such, they have been of great interest to researchers studying the endosymbiotic origins of organelles.
Other important ecological groups are:
- Granofilosea, comprising several groups traditionally considered heliozoa such as Heliomonadida, Desmothoracida and Gymnosphaerida.[10]
- Phaeodaria, marine protozoa previously considered radiolarians.[11]
Ecology
As well as being highly diverse in morphology and physiology, Cercozoa also shows high ecological diversity.[12] The phylum Cercozoa includes many of the most abundant and ecologically significant protozoa in soil, marine and freshwater ecosystems.[8]
Soil-dwelling cercozoans are one of the dominant groups of free-living eukaryotic microorganisms found in temperate soils, accounting for around 30% of identifiable protozoan DNA in arid or semi-arid soils and 15% in more humid soils. In transcriptomic analyses they account for 40-60% of all identifiable protozoan RNA found in forest and grassland soils. They also comprise 9-24% of all operational taxonomic units found in the ocean floor.[12]
Some cercozoa are coprophilic or coprozoic, meaning they use feces as a source of nutrients or as transport through animal hosts. The faecal habitat is an understudied reservoir of microbial eukaryotic diversity, dominated by amoeboflagellates from the phylum Cercozoa. Strongly coprophilic examples of cercozoa are the flagellates Cercomonas, Proleptomonas and Helkesimastix, and the sorocarpic amoeba Guttulinopsis. Many new cercozoan lineages, especially among sarcomonads, have been discovered through phylogenetic sampling of feces because they appear preferentially in this medium.[13]
Cercozoan bacterivores (i.e. predators of bacteria) are highly diverse and important in the plant phyllosphere, the leaf surfaces of plants. Particularly sarcomonads, with their ability to cyst, feed and multiply within hours, are perfectly adapted to the fluctuating environmental factors in the phyllosphere. Their predation causes shifts in the bacterial communities: they reduce populations of alphaproteobacteria and betaproteobacteria, which are less resistant to their grazing, in favour of other bacterial populations such as gammaproteobacteria.[14]
Evolution
External evolution
| Paraphyletic Cercozoa[2] | |||||||||||||||||||||||||||||||||
|
| Monophyletic Cercozoa[15] | |||||||||||||||||||||||||||||||||||||||
|
Originally, Cercozoa contained both Filosa and Endomyxa, according to phylogenetic analyses using ribosomal RNA and tubulin. These analyses also confirmed Cercozoa as the sister group of Retaria within the supergroup Rhizaria.[10][16]
However, the monophyly of the group was still uncertain. Posterior multigene phylogenetic analyses consistently found Cercozoa to be paraphyletic, because Endomyxa clustered next to Retaria instead of Filosa.[17][18][19] Because of this, Endomyxa was excluded from Cercozoa, which became a synonym of Filosa.[2]
More recent phylogenomic analyses with better sampling recovered a sister relationship between Filosa (=Cercozoa) and Endomyxa once again,[15] although the modern classification of eukaryotes retains Endomyxa, Cercozoa and Retaria as separate phyla within Rhizaria.[20]
Internal evolution
The phylum Cercozoa previously contained both Filosa and Endomyxa, but in the latest classifications Endomyxa has been excluded, and Cercozoa is now synonymous with Filosa. It is composed of two subphyla: Monadofilosa and Reticulofilosa. According to multigene phylogenetic analyses, Monadofilosa is a robust clade, in which the deepest branching group is Metromonadea, followed by Helkesea as the second group (together forming the paraphyletic Eoglissa) before the divergence of the clade Ventrifilosa (Imbricatea, Sarcomonadea and Thecofilosea). On the other hand, Reticulofilosa is probably paraphyletic, with Granofilosea diverging earlier than Chlorarachnea, which makes Chlorarachnea the sister group of Monadofilosa.[2]
| Cercozoa |
| |||||||||||||||||||||||||||||||||
A more recent phylogenomic analysis recovered both Monadofilosa and Reticulofilosa as monophyletic within the clade Filosa.[15]
In addition to the known Granofilosea, Chlorarachnea and Monadofilosa, a variety of clades inside Cercozoa have been discovered in other analyses and have slowly been described and named, such as Tremulida (previously known as Novel Clade 11)[16] and Aquavolonida (Novel Clade 10),[21] although their specific positions among the two main cercozoan subphyla have yet to be refined. These two orders have been classified as the class Skiomonadea, within Reticulofilosa.[2]
Classification
The classification of Cercozoa was revised in 2018:[2]
- Subphylum Reticulofilosa Cavaler-Smith 1997
- Class Chlorarachnea Hibberd & Norris 1984 (as Chlorarachniophyceae)
- Class Granofilosea Cavalier-Smith & Bass 2009
- Class Skiomonadea Cavalier-Smith 2012
- Subphylum Monadofilosa Cavalier-Smith 1997
- Superclass Eoglissa Cavalier-Smith 2011 emend. 2018
- Class Metromonadea Cavalier-Smith 2007
- Class Helkesea Cavalier-Smith 2018
- Superclass Ventrifilosa Cavalier-Smith 2012 emend. 2018
- Class Sarcomonadea Cavalier-Smith 1993 emend. 2018
- Class Imbricatea Cavalier-Smith 2003 emend. 2018
- Class Thecofilosea Cavalier-Smith 2003 emend. 2012
- Superclass Eoglissa Cavalier-Smith 2011 emend. 2018
Gallery
- Cercomonas sp. (Cercozoa: Cercomonadida)
 Ebria sp. (Cercozoa: Ebridea)
Ebria sp. (Cercozoa: Ebridea) Rhipidodendron sp. (Cercozoa: Spongomonadea)
Rhipidodendron sp. (Cercozoa: Spongomonadea) Euglypha sp. (Cercozoa: Euglyphida)
Euglypha sp. (Cercozoa: Euglyphida) Phaeodarians (Cercozoa: Phaeodarea)
Phaeodarians (Cercozoa: Phaeodarea) Clathrulina elegans (Cercozoa: Desmothoracida)
Clathrulina elegans (Cercozoa: Desmothoracida) Chlorarachnion sp. (Cercozoa: (Chlorarachniophyta)
Chlorarachnion sp. (Cercozoa: (Chlorarachniophyta) Vampyrella sp. (Cercozoa: Vampyrellidae)
Vampyrella sp. (Cercozoa: Vampyrellidae)
_b_083.jpg.webp)
 Auranticordis (Cercozoa: Marimonadida)
Auranticordis (Cercozoa: Marimonadida)
References
- Cavalier-Smith, T. (1998). "A revised six-kingdom system of life". Biological Reviews of the Cambridge Philosophical Society. 73 (3): 203–266. doi:10.1111/j.1469-185X.1998.tb00030.x. PMID 9809012. S2CID 6557779.
- Cavalier-Smith, Thomas; E. Chao, Ema; Lewis, Rhodri (2018), "Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria", Protoplasma, 255 (5): 1517–1574, doi:10.1007/s00709-018-1241-1, PMC 6133090, PMID 29666938
- Cavalier-Smith T (1997). "Amoeboflagellates and mitochondrial cristae in eukaryote evolution: megasystematics of the new protozoan subkingdoms eozoa and neozoa". Archiv für Protistenkunde. 147 (3–4): 237–258. doi:10.1016/S0003-9365(97)80051-6. ISSN 0003-9365.
- Nikolaev SI, Berney C, Fahrni JF, et al. (May 2004). "The twilight of Heliozoa and rise of Rhizaria, an emerging supergroup of amoeboid eukaryotes". Proc. Natl. Acad. Sci. U.S.A. 101 (21): 8066–71. doi:10.1073/pnas.0308602101. PMC 419558. PMID 15148395.
- Hoppenrath, M.; Leander B.S. (2006). "Ebriid phylogeny and the expansion of the Cercozoa". Protist. 157 (3): 279–90. doi:10.1016/j.protis.2006.03.002. PMID 16730229.
- Chantangsi, C. (2009). Comparative morphology and molecular evolution of marine interstitial cercozoans. PhD thesis. University of British Columbia.
- "SYSTEMATIC BIOLOGY: CERCOZOA". Retrieved 2009-03-28.
- Bass D, Cavalier-Smith T (1 November 2004). "Phylum-specific environmental DNA analysis reveals remarkably high global biodiversity of Cercozoa (Protozoa)". International Journal of Systematic and Evolutionary Microbiology. 54 (6): 2393–2404. doi:10.1099/ijs.0.63229-0. PMID 15545489.
- Cavalier-Smith T, Chao EE (October 2003). "Phylogeny and classification of phylum Cercozoa (Protozoa)" (PDF). Protist. 154 (3–4): 341–58. doi:10.1078/143446103322454112. PMID 14658494.
- Bass D, Chao EE, Nikolaev S, et al. (February 2009). "Phylogeny of Novel Naked Filose and Reticulose Cercozoa: Granofilosea cl. n. and Proteomyxidea Revised". Protist. 160 (1): 75–109. doi:10.1016/j.protis.2008.07.002. PMID 18952499.
- Nakamura, Yasuhide; Imai, Ichiro; Yamaguchi, Atsushi; Tuji, Akihiro; Not, Fabrice; Suzuki, Noritoshi (2015). "Molecular Phylogeny of the Widely Distributed Marine Protists, Phaeodaria (Rhizaria, Cercozoa)". Protist. 166 (3): 363–373. doi:10.1016/j.protis.2015.05.004. PMID 26083083.
- Harder C, Rønn R, Brejnrod A, et al. (8 March 2016). "Local diversity of heathland Cercozoa explored by in-depth sequencing". The ISME Journal. 10 (10): 2488–2497. doi:10.1038/ismej.2016.31. PMC 5030685. PMID 26953604.
- Bass D, Silberman JD, Brown MW, Pearce RA, Tice AK, Jousset A, Geisen S, Hartikainen H (23 February 2016). "Coprophilic amoebae and flagellates, including Guttulinopsis, Rosculus and Helkesimastix, characterise a divergent and diverse rhizarian radiation and contribute to a large diversity of faecal-associated protists". Environmental Microbiology. 18 (5): 1604–1619. doi:10.1111/1462-2920.13235. PMID 26914587.
- Flues S, Bass D, Bonkowski M (15 June 2017). "Grazing of leaf-associated Cercomonads (Protists: Rhizaria: Cercozoa) structures bacterial community composition and function". Environmental Microbiology. 19 (8): 3297–3309. doi:10.1111/1462-2920.13824. PMID 28618206.
- Irwin, Nicholas A.T.; Tikhonenkov, Denis V.; Hehenberger, Elisabeth; Mylnikov, Alexander P.; Burki, Fabien; Keeling, Patrick J. (2019-01-01). "Phylogenomics supports the monophyly of the Cercozoa". Molecular Phylogenetics and Evolution. 130: 416–423. doi:10.1016/j.ympev.2018.09.004. ISSN 1055-7903. PMID 30318266. S2CID 52982396.
- Howe, Alexis T.; Bass, David; Scoble, Josephine M.; Lewis, Rhodri; Vickerman, Keith; Arndt, Hartmut; Cavalier-Smith, Thomas (2011). "Novel Cultured Protists Identify Deep-branching Environmental DNA Clades of Cercozoa: New Genera Tremula, Micrometopion, Minimassisteria, Nudifila, Peregrinia". Protist. 162 (2): 332–372. doi:10.1016/j.protis.2010.10.002. ISSN 1434-4610. PMID 21295519.
- Burki F, Corradi N, Sierra R, Meyer GR, Abbott CL, Keeling PJ, et al. (July 2013). "Phylogenomics of the Intracellular Parasite Mikrocytos mackini Reveals Evidence for a Mitosome in Rhizaria". Current Biology. 23 (16): 1541–1547. doi:10.1016/j.cub.2013.06.033. PMID 23891116. S2CID 8257631.
- Burki F, Kudryavtsev A, Matz MV, et al. (2010). "Evolution of Rhizaria: new insights from phylogenomic analysis of uncultivated protists". BMC Evol Biol. 10: 377. doi:10.1186/1471-2148-10-377. PMC 3014934. PMID 21126361.
- Krabberød, Anders K.; Orr, Russell J.S.; Bråte, Jon; Kristensen, Tom; Bjørklund, Kjell R.; Shalchian-Tabrizi, Kamran (2017). "Single Cell Transcriptomics, Mega-Phylogeny, and the Genetic Basis of Morphological Innovations in Rhizaria". Mol. Biol. Evol. 34 (7): 1557–1573. doi:10.1093/molbev/msx075. PMC 5455982. PMID 28333264.
- Adl SM, Bass D, Lane CE, Lukeš J, Schoch CL, Smirnov A, Agatha S, Berney C, Brown MW, Burki F, Cárdenas P, Čepička I, Chistyakova L, del Campo J, Dunthorn M, Edvardsen B, Eglit Y, Guillou L, Hampl V, Heiss AA, Hoppenrath M, James TY, Karnkowska A, Karpov S, Kim E, Kolisko M, Kudryavtsev A, Lahr DJG, Lara E, Le Gall L, Lynn DH, Mann DG, Massana R, Mitchell EAD, Morrow C, Park JS, Pawlowski JW, Powell MJ, Richter DJ, Rueckert S, Shadwick L, Shimano S, Spiegel FW, Torruella G, Youssef N, Zlatogursky V, Zhang Q (2019). "Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes". Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi:10.1111/jeu.12691. PMC 6492006. PMID 30257078.
- Bass D, Tikhonenkov DV, Foster R, Dyal P, Janouškovec J, Keeling PJ, Gardner M, Neuhauser S, Hartikainen H, Mylnikov AP, Berney C (2018). "Rhizarian 'Novel Clade 10' Revealed as Abundant and Diverse Planktonic and Terrestrial Flagellates, including Aquavolon n. gen". Journal of Eukaryotic Microbiology. 65 (6): 828–842. doi:10.1111/jeu.12524. PMC 6282753. PMID 29658156.
External links
- Tree of Life Cercozoa
- phylogeny of Phaeodarea