Chromista
Chromista is a proposed but polyphyletic[1][2][3] biological kingdom consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids).[4] It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.[1]
| Chromista | |
|---|---|
 | |
| Chromista collage | |
| Scientific classification | |
| Domain: | Eukaryota |
| Clade: | Diaphoretickes |
| Kingdom: | Chromista Cavalier-Smith 1981 |
| Phyla | |
Chromista as a taxon was created by the British biologist Thomas Cavalier-Smith in 1981 to distinguish the stramenopiles, haptophytes, and cryptophytes.[5] According to Cavalier-Smith, the kingdom originally consisted mostly of photosynthetic eukaryotes (algae), but he later brought many heterotrophs (protozoa) into the proposed group. As of 2018, the kingdom was nearly as diverse as the Kingdoms Plantae and Animalia, consisting of eight phyla. Notable members include marine algae, potato blight, dinoflagellates, Paramecium, the brain parasite Toxoplasma, and the malarial parasite Plasmodium.[6]
However, Cavalier-Smith's hypothesis of chromist monophyly has been rejected by other researchers, who consider it more likely that some chromists acquired their plastids by incorporating another chromist instead of inheriting them from a common ancestor. This is thought to have occurred repeatedly, so that the red plastids spread from one group to another. The plastids, far from characterising their hosts as belonging to a single clade, thus have a different history from their disparate hosts. They appear to have originated in the Rhodophytina, and to have been transmitted to the Cryptophytina and from them to both the Ochrophyta and the Haptophyta, and then from these last to the Myzozoa.[3]
Biology
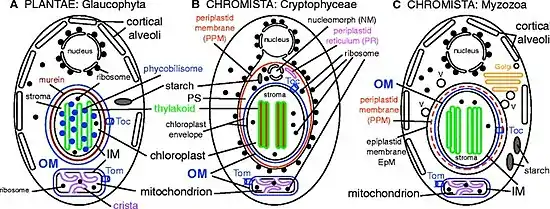
Members of Chromista are single-celled and multicellular eukaryotes having basically either or both features:[5]
- plastid(s) that contain chlorophyll c and lie within an extra (periplastid) membrane in the lumen of the rough endoplasmic reticulum (typically within the perinuclear cisterna);
- cilia with tripartite or bipartite rigid tubular hairs.
The kingdom includes diverse organisms from algae to malarial parasites (Plasmodium).[7] Molecular evidence indicates that the plastids in chromists were derived from red algae through secondary symbiogenesis in a single event.[8] In contrast, plants acquired their plastids from cyanobacteria through primary symbiogenesis.[9] These plastids are now enclosed in two extra cell membranes, making a four-membrane envelope, as a result of which they acquired many other membrane proteins for transporting molecules in and out of the organelles. The diversity of chromists is hypothesised to have arisen from degeneration, loss or replacement of the plastids in some lineages.[10] Additional symbiogenesis of green algae has provided genes retained in some members (such as heterokonts),[11] and bacterial chlorophyll (indicated by the presence of ribosomal protein L36 gene, rpl36) in haptophytes and cryptophytes.[12]
History and groups
Some examples of classification of the groups involved, which have overlapping but non-identical memberships, are shown below.[13][14]
Chromophycées (Chadefaud, 1950)
The Chromophycées (Chadefaud, 1950),[15] renamed Chromophycota (Chadefaud, 1960),[16] included the current Ochrophyta (autotrophic Stramenopiles), Haptophyta (included in Chrysophyceae until Christensen, 1962), Cryptophyta, Dinophyta, Euglenophyceae and Choanoflagellida (included in Chrysophyceae until Hibberd, 1975).
Chromophyta (Christensen 1962, 1989)
The Chromophyta (Christensen 1962, 2008), defined as algae with chlorophyll c, included the current Ochrophyta (autotrophic Stramenopiles), Haptophyta, Cryptophyta, Dinophyta and Choanoflagellida. The Euglenophyceae were transferred to the Chlorophyta.
Chromophyta (Bourrelly, 1968)
The Chromophyta (Bourrelly, 1968) included the current Ochrophyta (autotrophic Stramenopiles), Haptophyta and Choanoflagellida. The Cryptophyceae and the Dinophyceae were part of Pyrrhophyta (= Dinophyta).
Chromista (Cavalier-Smith, 1981)
The name Chromista was first introduced by Cavalier-Smith in 1981;[5] the earlier names Chromophyta, Chromobiota and Chromobionta correspond to roughly the same group. It has been described as consisting of three different groups:[17] It includes all protists whose plastids contain chlorophyll c.[1]
- Heterokonts or Stramenopiles: brown algae, diatoms, water moulds, etc.
- Haptophytes
- Cryptomonads
In 1994, Cavalier-Smith and colleagues indicated that the Chromista is probably a polyphyletic group whose members arose independently, sharing no more than descent from the common ancestor of all eukaryotes:[1]
The four phyla that contain chromophyte algae (those with chlorophyll c--i.e., Cryptista, Heterokonta, Haptophyta, Dinozoa) are distantly related to each other and to Chlorarachniophyta on our trees. However, all of the photosynthetic taxa within each of these four phyla radiate from each other very substantially after the radiation of the four phyla themselves. This favors the view that the common ancestor of these four phyla was not photosynthetic and that chloroplasts were implanted separately into each much more recently. This probable polyphyly of the chromophyte algae, if confirmed, would make it desirable to treat Cryptista, Heterokonta, and Haptophyta as separate kingdoms, rather than to group them together in the single kingdom Chromista.[1]
In 2009, Cavalier-Smith gave his reason for making a new kingdom, saying:[7]
I established Chromista as a kingdom distinct from Plantae and Protozoa because of the evidence that chromist chloroplasts were acquired secondarily by enslavement of a red alga, itself a member of kingdom Plantae, and their unique membrane topology.[7]
Since then Chromista has been defined in different ways at different times. In 2010, Cavalier-Smith reorganised Chromista to include the SAR supergroup (named for the included groups Stramenopiles, Alveolata and Rhizaria) and Hacrobia (Haptista and Cryptista).[7]
Patron et al. (2004) considered the presence of a unique class of FBA (fructose-1,6-biophosphate-aldolase) enzyme not similar to that found in plants as evidence of chromist monophyly.[18] Fast et al. (2001) supported a single origin for the myzozoan (dinoflagellate + apicomplexan), heterokont and cryptophyte plastids based on their comparison of GAPDH (glyceraldehyde-3-phosphate dehydrogenase) genes.[19] Harper & Keeling (2003) described haptophyte homologs and considered them further evidence of a single endosymbiotic event involving the ancestor of all chromists.[20]
Chromalveolata (Adl et al., 2005)
The Chromalveolata included Stramenopiles, Haptophyta, Cryptophyta and Alveolata.[21] However, in 2008 the group was found not to be monophyletic,[22][23] and later studies confirmed this.[24][25]
Classification
Cavalier-Smith et al. 2015
In 2015, Cavalier-Smith and his colleagues made a new higher-level grouping of all organisms as a revision of the seven kingdoms model. In it, they classified the kingdom Chromista into 2 subkingdoms and 11 phyla, namely:[26]
| Chromista |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2015 |
Cavalier-Smith 2018
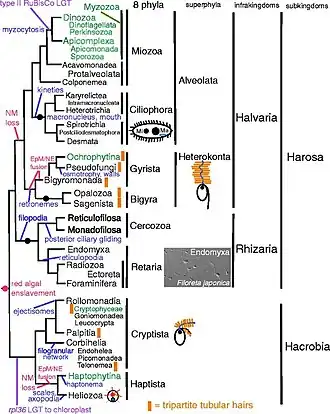
Cavalier-Smith made a new analysis of Chromista in 2018 in which he classified all chromists into 8 phyla (Gyrista corresponds to the above phyla Ochrophyta and Pseudofungi, Cryptista corresponds to the above phyla Cryptista and "N.N.", Haptista corresponds to the above phyla Haptophyta and Heliozoa):[6]
| Chromista |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| 2018 |
Polyphyly and serial endosymbiosis
Molecular trees have had difficulty resolving relationships between the different groups. All three may share a common ancestor with the alveolates (see chromalveolates), but there is evidence that suggests the haptophytes and cryptomonads do not belong together with the heterokonts or the SAR clade, but may be associated with the Archaeplastida.[4][27] Cryptista specifically may be sister or part of Archaeplastida,[24] though this could be an artefact due to acquisition of genes from red algae by cryptomonads.[3]
A 2020 phylogeny of the eukaryotes states that "the chromalveolate hypothesis is not widely accepted" (noting Cavalier-Smith et al 2018[28] as an exception), explaining that the host lineages do not appear to be closely related in "most phylogenetic analyses".[2][29] Further, none of TSAR, Cryptista, and Haptista, groups formerly within Chromalveolata, appear "likely to be ancestrally defined by red secondary plastids". This is because of the many non-photosynthetic organisms related to the groups with chlorophyll c, and the possibility that cryptophytes are more closely related to plants.[2]
The alternative to monophyly is serial endosymbiosis, meaning that the "chromists" acquired their plastids from each other instead of inheriting them from a single common ancestor. Thus the phylogeny of the distinctive plastids, which are agreed to have a common origin in the rhodophytes, is different from the phylogeny of the host cells.[3] In 2021, Jürgen Strassert and colleagues modelled the timelines for the presumed spread of the red plastids, concluding that "the hypotheses of serial endosymbiosis are chronologically possible, as the stem lineages of all red plastid-containing groups overlap in time" during the Mesoproterozoic and Neoproterozoic eras. They propose that the plastids were transmitted between groups as follows:[3]
References
- Cavalier-Smith, Thomas; Allsopp, M. T.; Chao, E. E. (November 1994). "Chimeric conundra: are nucleomorphs and chromists monophyletic or polyphyletic?". Proceedings of the National Academy of Sciences of the United States of America. 91 (24): 11368–11372. Bibcode:1994PNAS...9111368C. doi:10.1073/pnas.91.24.11368. PMC 45232. PMID 7972066.
- Burki, Fabien; Roger, Andrew J.; Brown, Matthew W.; Simpson, Alastair G.B. (2020). "The New Tree of Eukaryotes". Trends in Ecology & Evolution. Elsevier. 35 (1): 43–55. doi:10.1016/j.tree.2019.08.008. ISSN 0169-5347. PMID 31606140. S2CID 204545629.
- Strassert, Jürgen F. H.; Irisarri, Iker; Williams, Tom A.; Burki, Fabien (2021-03-25). "A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids". Nature Communications. 12 (1): 1879. Bibcode:2021NatCo..12.1879S. doi:10.1038/s41467-021-22044-z. ISSN 2041-1723. PMC 7994803. PMID 33767194.
- Parfrey, Laura Wegener; Barbero, Erika; Lasser, Elyse; et al. (December 2006). "Evaluating support for the current classification of eukaryotic diversity". PLOS Genetics. 2 (12): e220. doi:10.1371/journal.pgen.0020220. PMC 1713255. PMID 17194223.
- Cavalier-Smith, Thomas (1981). "Eukaryote kingdoms: seven or nine?". Bio Systems. 14 (3–4): 461–81. doi:10.1016/0303-2647(81)90050-2. PMID 7337818.
- Cavalier-Smith, Thomas (2018). "Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences". Protoplasma. 255 (1): 297–357. doi:10.1007/s00709-017-1147-3. PMC 5756292. PMID 28875267.
- Cavalier-Smith, Thomas (2009). "Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree". Biology Letters. 6 (3): 342–345. doi:10.1098/rsbl.2009.0948. PMC 2880060. PMID 20031978.
- Keeling, Patrick J. (2009). "Chromalveolates and the Evolution of Plastids by Secondary Endosymbiosis". Journal of Eukaryotic Microbiology. 56 (1): 1–8. doi:10.1111/j.1550-7408.2008.00371.x. PMID 19335769. S2CID 34259721.
- Ponce-Toledo, Rafael I.; Deschamps, Philippe; López-García, Purificación; Zivanovic, Yvan; Benzerara, Karim; Moreira, David (2017). "An Early-Branching Freshwater Cyanobacterium at the Origin of Plastids". Current Biology. 27 (3): 386–391. doi:10.1016/j.cub.2016.11.056. PMC 5650054. PMID 28132810.
- Keeling, Patrick J. (2010). "The endosymbiotic origin, diversification and fate of plastids". Philosophical Transactions of the Royal Society B: Biological Sciences. 365 (1541): 729–748. doi:10.1098/rstb.2009.0103. PMC 2817223. PMID 20124341.
- Morozov, A. A.; Galachyants, Yuri P. (2019). "Diatom genes originating from red and green algae: Implications for the secondary endosymbiosis models". Marine Genomics. 45: 72–78. Bibcode:2019MarGn..45...72M. doi:10.1016/j.margen.2019.02.003. PMID 30792089. S2CID 73458340.
- Rice, Danny W; Palmer, Jeffrey D (2006). "An exceptional horizontal gene transfer in plastids: gene replacement by a distant bacterial paralog and evidence that haptophyte and cryptophyte plastids are sisters". BMC Biology. 4 (1): 31. doi:10.1186/1741-7007-4-31. PMC 1570145. PMID 16956407.
- de Reviers, Bruno (2006). Biologia e Filogenia das Algas. Porto Alegre: Editora Artmed. pp. 156–157. ISBN 9788536315102.
- Blackwell, Will H. (2009). "Chromista revisited: a dilemma of overlapping putative kingdoms, and the attempted application of the botanical code of nomenclature" (PDF). Phytologia. 91 (2): 191–225.
- Chadefaud, Marius (1950). "Les cellules nageuses des Algues dans l'embranchement des Chromophycées" [Seaweed swimming cells in the branch of Chromophyceae]. Comptes rendus hebdomadaires des séances de l'Académie des Sciences (in French). 231: 788–790.
- Chadefaud, Marius (1960). "Les végétaux non vasculaires (Cryptogamie)". In Chadefaud, Marius; Emberger, L. (eds.). Traité de Botanique Systématique. Paris: Tome I.
- Csurös, M.; Rogozin, I. B.; Koonin, Eugene V. (May 2008). "Extremely intron-rich genes in the alveolate ancestors inferred with a flexible maximum-likelihood approach". Molecular Biology and Evolution. 25 (5): 903–911. doi:10.1093/molbev/msn039. PMID 18296415.
- Patron, Nicola J.; Rogers, Matthew B.; Keeling, Patrick J. (2004). "Gene Replacement of Fructose-1,6-Bisphosphate Aldolase Supports the Hypothesis of a Single Photosynthetic Ancestor of Chromalveolates". Eukaryotic Cell. 3 (5): 1169–1175. doi:10.1128/EC.3.5.1169-1175.2004. ISSN 1535-9778. PMC 522617. PMID 15470245.
- Fast, Naomi M.; Kissinger, Jessica C.; Roos, David S.; Keeling, Patrick J. (2001-03-01). "Nuclear-Encoded, Plastid-Targeted Genes Suggest a Single Common Origin for Apicomplexan and Dinoflagellate Plastids". Molecular Biology and Evolution. 18 (3): 418–426. doi:10.1093/oxfordjournals.molbev.a003818. ISSN 1537-1719. PMID 11230543.
- Harper, J. T. (2003-06-27). "Nucleus-Encoded, Plastid-Targeted Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) Indicates a Single Origin for Chromalveolate Plastids". Molecular Biology and Evolution. 20 (10): 1730–1735. doi:10.1093/molbev/msg195. ISSN 0737-4038. PMID 12885964.
- Adl, Sina M.; et al. (2005). "The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists". Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111/j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- Burki, Fabien; Shalchian-Tabrizi, Kamran & Pawlowski, Jan (2008). "Phylogenomics reveals a new 'megagroup' including most photosynthetic eukaryotes". Biology Letters. 4 (4): 366–369. doi:10.1098/rsbl.2008.0224. PMC 2610160. PMID 18522922.
- Kim, E.; Graham, L. E. (July 2008). Redfield, Rosemary Jeanne (ed.). "EEF2 analysis challenges the monophyly of Archaeplastida and Chromalveolata". PLOS ONE. 3 (7): e2621. Bibcode:2008PLoSO...3.2621K. doi:10.1371/journal.pone.0002621. PMC 2440802. PMID 18612431.
- Burki, Fabien; Kaplan, Maia; Tikhonenkov, Denis V.; et al. (January 2016). "Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista". Proceedings. Biological Sciences. 283 (1823): 20152802. doi:10.1098/rspb.2015.2802. PMC 4795036. PMID 26817772.
- Burki, F.; Okamoto, N.; Pombert, J. F.; Keeling, P. J. (2012). "The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins". Proceedings of the Royal Society. 279 (1736): 2246–2254. doi:10.1098/rspb.2011.2301. PMC 3321700. PMID 22298847.
- Ruggiero, Michael A.; Gordon, Dennis P.; Orrell, Thomas M.; et al. (2015). "A higher level classification of all living organisms". PLOS ONE. 10 (4): e0119248. Bibcode:2015PLoSO..1019248R. doi:10.1371/journal.pone.0119248. PMC 4418965. PMID 25923521.
- Burki, Fabien; Shalchian-Tabrizi, Kamran; Minge, Marianne; et al. (August 2007). "Phylogenomics reshuffles the eukaryotic supergroups". PLOS ONE. 2 (8): e790. Bibcode:2007PLoSO...2..790B. doi:10.1371/journal.pone.0000790. PMC 1949142. PMID 17726520.
- Cavalier-Smith, Thomas; Chao, Ema E.; Lewis, Rhodri (17 April 2018). "Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria". Protoplasma. 255 (5): 1517–1574. doi:10.1007/s00709-018-1241-1. ISSN 0033-183X. PMC 6133090. PMID 29666938.
- Burki, F. (2017). "The convoluted evolution of eukaryotes with complex plastids". In Hirakawa, Y. (ed.). Advances in Botanical Research. Vol. 84. Academic Press. pp. 1–30.