Springtail
Springtails (Collembola) form the largest of the three lineages of modern hexapods that are no longer considered insects (the other two are the Protura and Diplura). Although the three orders are sometimes grouped together in a class called Entognatha because they have internal mouthparts, they do not appear to be any more closely related to one another than they are to all insects, which have external mouthparts.
| Springtails Temporal range: | |
|---|---|
 | |
| Orchesella cincta | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Clade: | Pancrustacea |
| Subphylum: | Hexapoda |
| Subclass: | Collembola Lubbock, 1871 |
| Orders | |
| |
| Synonyms [1] | |
| |
Collembolans are omnivorous, free-living organisms that prefer moist conditions. They do not directly engage in the decomposition of organic matter, but contribute to it indirectly through the fragmentation of organic matter[2] and the control of soil microbial communities.[3] The word Collembola is from the ancient Greek κόλλα kólla "glue" and ἔμβολος émbolos "peg"; this name was given due to the existence of the collophore, which was previously thought to stick to surfaces to stabilize the creature.[4]
Some DNA sequence studies[5][6][7] suggest that Collembola represent a separate evolutionary line from the other Hexapoda, but others disagree;[8] this seems to be caused by widely divergent patterns of molecular evolution among the arthropods.[9] The adjustments of traditional taxonomic rank for springtails reflect the occasional incompatibility of traditional groupings with modern cladistics: when they were included with the insects, they were ranked as an order; as part of the Entognatha, they are ranked as a subclass. If they are considered a basal lineage of Hexapoda, they are elevated to full class status.
Morphology


Members of the Collembola are normally less than 6 mm (0.24 in) long, have six or fewer abdominal segments, and possess a tubular appendage (the collophore or ventral tube) with reversible, sticky vesicles, projecting ventrally from the first abdominal segment.[10] It is believed to be associated with fluid uptake and balance, excretion, and orientation of the organism itself.[11] Most species have an abdominal, tail-like appendage known as a furcula (or furca). It is located on the fourth abdominal segment of collembolans and is folded beneath the body, held under tension by a small structure called the retinaculum (or tenaculum). When released, it snaps against the substrate, flinging the springtail into the air and allowing for rapid evasion and travel. All of this takes place in as little as 18 milliseconds.[12][11]
Springtails also possess the ability to reduce their body size by as much as 30% through subsequent ecdyses (moulting) if temperatures rise high enough. The shrinkage is genetically controlled. Since warmer conditions increase metabolic rates and energy requirements in organisms, the reduction in body size is advantageous to their survival.[13]
The Poduromorpha and Entomobryomorpha have an elongated body, while the Symphypleona and Neelipleona have a globular body. Collembola lack a tracheal respiration system, which forces them to respire through a porous cuticle, except for the two families Sminthuridae and Actaletidae, which exhibit a single pair of spiracles between the head and the thorax, leading to a rudimentary, although fully functional, tracheal system.[14][10] The anatomical variance present between different species partially depends on soil morphology and composition. Surface-dwellers are generally larger, have darker pigments, have longer antennae and functioning furcula. Sub-surface-dwellers are usually unpigmented, have elongated bodies, and reduced furcula. They can be categorized into four main forms according to soil composition and depth: atmobiotic, epedaphic, hemiedaphic, and euedaphic. Atmobiotic species inhabit macrophytes and litter surfaces. They are generally 8-10 millimeters (about ⅓") in length, pigmented, have long limbs, and a full set of ocelli (photoreceptors). Epedaphic species inhabit upper litter layers and fallen logs. They are slightly smaller and have less pronounced pigments, as well as less developed limbs and ocelli than the atmobiotic species. Hemiedaphic species inhabit the lower litter layers of decomposing organic material. They are 1-2 millimeters (about 1/16") in length, have dispersed pigmentation, shortened limbs, and a reduced number of ocelli. Euedaphic species inhabit upper mineral layers known as the humus horizon. They are smaller than hemiedaphic species; have soft, elongated bodies; lack pigmentation and ocelli; and have reduced or absent furca.[15][16][17]
Poduromorphs are characterized by their elongated bodies and conspicuous segmentation – three thoracic segments, six abdominal segments, including a well-developed prothorax with tergal chaetae,[17] while the first thoracic segment in Entomobryomorpha is clearly reduced and bears no chaetae.
The digestive tract of collembolan species consists of three main components: the foregut, midgut, and hindgut. The midgut is surrounded by a network of muscles and lined with a monolayer of columnar or cuboidal cells. Its function is to mix and transport food from the lumen into the hindgut through contraction. Many species of syntrophic bacteria, archaea, and fungi are present in the lumen. These different digestive regions have varying pH to support specific enzymatic activities and microbial populations. The anterior portion of the midgut and hindgut is slightly acidic (with a pH of approximately 6.0) while the posterior midgut portion is slightly alkaline (with a pH of approximately 8.0). Between the midgut and hindgut is an alimentary canal called the pyloric region, which is a muscular sphincter.[11]
Systematics and evolution
Traditionally, the springtails were divided into the orders Arthropleona, Symphypleona, and occasionally also Neelipleona. The Arthropleona were divided into two superfamilies, the Entomobryoidea and the Poduroidea. However, recent phylogenetic studies show Arthropleona is paraphyletic.[18][19][20] Thus, the Arthropleona are abolished in modern classifications, and their superfamilies are raised in rank accordingly, being now orders Entomobryomorpha and the Poduromorpha. Technically, the Arthropleona are thus a partial junior synonym of the Collembola.[21]
The term "Neopleona" is essentially synonymous with Symphypleona + Neelipleona.[22] The Neelipleona was originally seen as a particularly advanced lineage of Symphypleona, based on the shared global body shape, but the global body of the Neelipleona is realized in a completely different way than in Symphypleona. Subsequently, the Neelipleona were considered as being derived from the Entomobryomorpha. Analysis of 18S and 28S rRNA sequence data, though, suggests that they form the most ancient lineage of springtails, which would explain their peculiar apomorphies.[8] This phylogenetic relationship was also confirmed using a phylogeny based on mtDNA[19] and whole-genome data.[20]
The latest whole-genome phylogeny supporting four orders of Collembola:[20]
| |||||||||||||||||||

Springtails are attested to since the Early Devonian.[23] The fossil from 400 million years ago, Rhyniella praecursor, is the oldest terrestrial arthropod, and was found in the famous Rhynie chert of Scotland. Given its morphology resembles extant species quite closely, the radiation of the Hexapoda can be situated in the Silurian, 420 million years ago or more.[24] Additional research concerning the coprolites (fossilized feces) of ancient collembolans allowed researchers to track their lineages back some 412 million years.[11]
Fossil Collembola are rare. Instead, most are found in amber.[25] Even these are rare and many amber deposits carry few or no collembola. The best deposits are from the early Eocene of Canada and Europe,[26] Miocene of Central America,[27] and the mid-Cretaceous of Burma and Canada.[28] They display some unexplained characteristics: first, all but one of the fossils from the Cretaceous belong to extinct genera, whereas none of the specimens from the Eocene or the Miocene are of extinct genera; second, the species from Burma are more similar to the modern fauna of Canada than are the Canadian Cretaceous specimens.
There are about 3,600 different species.[29]
Ecology
Eating behavior
Specific feeding strategies and mechanisms are employed to match specific niches.[30] Herbivorous and detritivorous species fragment biological material present in soil and leaf litter, supporting decomposition and increasing the availability of nutrients for various species of microbes and fungi.[31] Carnivorous species maintain populations of small invertebrates such as nematodes, rotifers, and other collembolan species.[11][15] Springtails commonly consume fungal hyphae and spores, but also have been found to consume plant material and pollen, animal remains, colloidal materials, minerals and bacteria.[32]
Predators
Springtails are consumed by mesostigmatan mites in various families, including Ascidae, Laelapidae, Parasitidae, Rhodacaridae and Veigaiidae.[33]
Cave-dwelling springtails are a food source for spiders and harvestmen in the same environment, such as the endangered harvestman Texella reyesi.[34]
Distribution
Springtails are cryptozoa frequently found in leaf litter and other decaying material,[35] where they are primarily detritivores and microbivores, and one of the main biological agents responsible for the control and the dissemination of soil microorganisms.[36] In a mature deciduous woodland in temperate climate, leaf litter and vegetation typically support 30 to 40 species of springtails, and in the tropics the number may be over 100.[37]

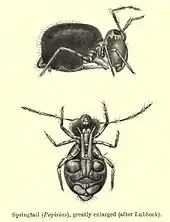
In sheer numbers, they are reputed to be one of the most abundant of all macroscopic animals, with estimates of 100,000 individuals per square meter of ground,[38] essentially everywhere on Earth where soil and related habitats (moss cushions, fallen wood, grass tufts, ant and termite nests) occur.[39] Only nematodes, crustaceans, and mites are likely to have global populations of similar magnitude, and each of those groups except mites is more inclusive. Though taxonomic rank cannot be used for absolute comparisons, it is notable that nematodes are a phylum and crustaceans a subphylum. Most springtails are small and difficult to see by casual observation, but one springtail, the so-called snow flea (Hypogastrura nivicola), is readily observed on warm winter days when it is active and its dark color contrasts sharply with a background of snow.[40]
In addition, a few species routinely climb trees and form a dominant component of canopy fauna, where they may be collected by beating or insecticide fogging.[41][42] These tend to be the larger (>2 mm) species, mainly in the genera Entomobrya and Orchesella, though the densities on a per square meter basis are typically 1–2 orders of magnitude lower than soil populations of the same species. In temperate regions, a few species (e.g. Anurophorus spp., Entomobrya albocincta, Xenylla xavieri, Hypogastrura arborea) are almost exclusively arboreal.[39] In tropical regions a single square meter of canopy habitat can support many species of Collembola.[12]
The main ecological factor driving the local distribution of species is the vertical stratification of the environment: in woodland a continuous change in species assemblages can be observed from tree canopies to ground vegetation then to plant litter down to deeper soil horizons.[39] This is a complex factor embracing both nutritional and physiological requirements, together with behavioural trends,[43] dispersal limitation[44] and probable species interactions. Some species have been shown to exhibit negative[45] or positive[43] gravitropism, which adds a behavioural dimension to this still poorly understood vertical segregation. Experiments with peat samples turned upside down showed two types of responses to disturbance of this vertical gradient, called "stayers" and "movers".[46]

As a group, springtails are highly sensitive to desiccation, because of their tegumentary respiration,[47] although some species with thin, permeable cuticles have been shown to resist severe drought by regulating the osmotic pressure of their body fluid.[48] The gregarious behaviour of Collembola, mostly driven by the attractive power of pheromones excreted by adults,[49] gives more chance to every juvenile or adult individual to find suitable, better protected places, where desiccation could be avoided and reproduction and survival rates (thereby fitness) could be kept at an optimum.[50] Sensitivity to drought varies from species to species[51] and increases during ecdysis.[52] Given that springtails moult repeatedly during their entire life (an ancestral character in Hexapoda) they spend much time in concealed micro-sites where they can find protection against desiccation and predation during ecdysis, an advantage reinforced by synchronized moulting.[53] The high humidity environment of many caves also favours springtails and there are numerous cave adapted species,[54][55] including one, Plutomurus ortobalaganensis living 1,980 metres (6,500 ft) down the Krubera Cave.[56]

The horizontal distribution of springtail species is affected by environmental factors which act at the landscape scale, such as soil acidity, moisture and light.[39] Requirements for pH can be reconstructed experimentally.[57] Altitudinal changes in species distribution can be at least partly explained by increased acidity at higher elevation.[58] Moisture requirements, among other ecological and behavioural factors, explain why some species cannot live aboveground,[59] or retreat in the soil during dry seasons,[60] but also why some epigeal springtails are always found in the vicinity of ponds and lakes, such as the hygrophilous Isotomurus palustris.[61] Adaptive features, such as the presence of a fan-like wettable mucro, allow some species to move at the surface of water (Sminthurides aquaticus, Sminthurides malmgreni). Podura aquatica, a unique representative of the family Poduridae (and one of the first springtails to have been described by Carl Linnaeus), spends its entire life at the surface of water, its wettable eggs dropping in water until the non-wettable first instar hatches then surfaces.[62]
In a variegated landscape, made of a patchwork of closed (woodland) and open (meadows, cereal crops) environments, most soil-dwelling species are not specialized and can be found everywhere, but most epigeal and litter-dwelling species are attracted to a particular environment, either forested or not.[39][63] As a consequence of dispersal limitation, landuse change, when too rapid, may cause the local disappearance of slow-moving, specialist species,[64] a phenomenon the measure of which has been called colonisation credit.[65][66]
Relationship with humans

Springtails are well known as pests of some agricultural crops. Sminthurus viridis, the lucerne flea, has been shown to cause severe damage to agricultural crops,[67] and is considered as a pest in Australia.[68][69] Onychiuridae are also known to feed on tubers and to damage them to some extent.[70] However, by their capacity to carry spores of mycorrhizal fungi and mycorrhiza helper bacteria on their tegument, soil springtails play a positive role in the establishment of plant-fungal symbioses and thus are beneficial to agriculture.[71] They also contribute to controlling plant fungal diseases through their active consumption of mycelia and spores of damping-off and pathogenic fungi.[72][73] It has been suggested that they could be reared to be used for the control of pathogenic fungi in greenhouses and other indoor cultures.[74][75]
Various sources and publications have suggested that some springtails may parasitize humans, but this is entirely inconsistent with their biology, and no such phenomenon has ever been scientifically confirmed, though it has been documented that the scales or hairs from collembolans can cause irritation when rubbed onto the skin.[76] They may sometimes be abundant indoors in damp places such as bathrooms and basements, and incidentally found on one's person. More often, claims of persistent human skin infection by springtails may indicate a neurological problem, such as delusional parasitosis, a psychological rather than entomological problem. Researchers themselves may be subject to psychological phenomena. For example, a publication in 2004 claiming that springtails had been found in skin samples was later determined to be a case of pareidolia; that is, no springtail specimens were actually recovered, but the researchers had digitally enhanced photos of sample debris to create images resembling small arthropod heads, which then were claimed to be springtail remnants.[76][77][78][79][80] However, Steve Hopkin reports one instance of an entomologist aspirating an Isotoma species and in the process accidentally inhaling some of their eggs, which hatched in his nasal cavity and made him quite ill until they were flushed out.[37]
In 1952, China accused the United States military of spreading bacteria-laden insects and other objects during the Korean War by dropping them from P-51 fighters above rebel villages over North Korea. In all, the U.S. was accused of dropping ants, beetles, crickets, fleas, flies, grasshoppers, lice, springtails, and stoneflies as part of a biological warfare effort. The alleged associated diseases included anthrax, cholera, dysentery, fowl septicemia, paratyphoid, plague, scrub typhus, small pox, and typhoid. China created an international scientific commission for investigating possible bacterial warfare, eventually ruling that the United States probably did engage in limited biological warfare in Korea. The US government denied all the allegations, and instead proposed that the United Nations send a formal inquiry committee to China and Korea, but China and Korea refused to cooperate. U.S. and Canadian entomologists further claimed that the accusations were ridiculous and argued that anomalous appearances of insects could be explained through natural phenomena.[81] Springtail species cited in allegations of biological warfare in the Korean War were Isotoma (Desoria) negishina (a local species) and the "white rat springtail" Folsomia candida.[82]
Captive springtails are often kept in a terrarium as part of a clean-up crew.[83]
Ecotoxicology laboratory animals
Springtails are currently used in laboratory tests for the early detection of soil pollution. Acute and chronic toxicity tests have been performed by researchers, mostly using the parthenogenetic isotomid Folsomia candida.[84] These tests have been standardized.[85] Details on a ringtest, on the biology and ecotoxicology of Folsomia candida and comparison with the sexual nearby species Folsomia fimetaria (sometimes preferred to Folsomia candida) are given in a document written by Paul Henning Krogh.[86] Care should be taken that different strains of the same species may be conducive to different results. Avoidance tests have been also performed.[87] They have been standardized, too.[88] Avoidance tests are complementary to toxicity tests, but they also offer several advantages: they are more rapid (thus cheaper), more sensitive and they are environmentally more reliable, because in the real world Collembola move actively far from pollution spots.[89] It may be hypothesized that the soil could become locally depauperated in animals (and thus improper to normal use) while below thresholds of toxicity. Contrary to earthworms, and like many insects and molluscs, Collembola are very sensitive to herbicides and thus are threatened in no-tillage agriculture, which makes a more intense use of herbicides than conventional agriculture.[90] The springtail Folsomia candida is also becoming a genomic model organism for soil toxicology.[91][92] With microarray technology the expression of thousands of genes can be measured in parallel. The gene expression profiles of Folsomia candida exposed to environmental toxicants allow fast and sensitive detection of pollution, and additionally clarifies molecular mechanisms causing toxicology.
Collembola have been found to be useful as bio-indicators of soil quality. Laboratory studies have been conducted that validated that the jumping ability of springtails can be used to evaluate the soil quality of Cu- and Ni-polluted sites.[93]
Climate warming impact
In polar regions that are expected to experience among the most rapid impact from climate warming, springtails have shown contrasting responses to warming in experimental warming studies.[94] There are negative,[95][96] positive[97][98] and neutral responses reported.[96][99] Neutral responses to experimental warming have also been reported in studies of non-polar regions.[100] The importance of soil moisture has been demonstrated in experiments using infrared heating in an alpine meadow, which had a negative effect on mesofauna biomass and diversity in drier parts and a positive effect in moist sub-areas.[101] Furthermore, a study with 20 years of experimental warming in three contrasting plant communities found that small scale heterogeneity may buffer springtails to potential climate warming.[99]
Reproduction
Sexual reproduction occurs through the clustered or scattered deposition of spermatophores by male adults. Stimulation of spermatophore deposition by female pheromones has been demonstrated in Sinella curviseta.[102] Mating behavior can be observed in Symphypleona.[103] Among Symphypleona, males of some Sminthuridae use a clasping organ located on their antenna.[35] Many collembolan species, mostly those living in deeper soil horizons, are parthenogenetic, which favors reproduction to the detriment of genetic diversity and thereby to population tolerance of environmental hazards.[104] Parthenogenesis (also called thelytoky) is under the control of symbiotic bacteria of the genus Wolbachia, which live, reproduce and are carried in female reproductive organs and eggs of Collembola.[105] Feminizing Wolbachia species are widespread in arthropods[106] and nematodes,[107] where they co-evolved with most of their lineages.
See also
References
- Gillott, Cedric (2005). "Apterygote hexapods". Entomology (3rd ed.). Berlin: Springer. pp. 113–125. doi:10.1007/1-4020-3183-1_5. ISBN 978-0-306-44967-3.
- Brady, Nyle C. & Weil, Ray R. (2009). "Organisms and ecology of the soil". Elements of the nature and properties of soils (3rd ed.). Upper Saddle River: Prentice Hall. ISBN 978-0-13-501433-2. OCLC 276340542.
- Thimm, Torsten; Hoffmann, Andrea; Borkott, Heinz; Munch, Jean Charles & Tebbe, Christoph C. (1998). "The gut of the soil microarthropod Folsomia candida (Collembola) is a frequently changeable but selective habitat and a vector for microorganisms". Applied and Environmental Microbiology. 64 (7): 2660–2669. Bibcode:1998ApEnM..64.2660T. doi:10.1128/AEM.64.7.2660-2669.1998. PMC 106441. PMID 9647845.
- Lubbock, John (1873). Monograph of the Collembola and Thysanura. London: Ray Society. p. 36.
- Nardi, Francesco; Spinsanti, Giacomo; Boore, Jeffrey L.; Carapelli, Antonio; Dallai, Romano & Frati, Francesco (2003). "Hexapod origins: monophyletic or paraphyletic?" (PDF). Science. 299 (5614): 1887–1889. Bibcode:2003Sci...299.1887N. doi:10.1126/science.1078607. PMID 12649480. S2CID 38792657.
- Delsuc, Frédéric; Phillips, Matthew J. & Penny, David (2003). "Comment on Hexapod origins: monophyletic or paraphyletic?" (PDF). Science. 301 (5639): 1482. doi:10.1126/science.1086558. PMID 12970547. S2CID 43942720.
- Nardi, Francesco; Spinsanti, Giacomo; Boore, Jeffrey L.; Carapelli, Antonio; Dallai, Romano & Frati, Francesco (2003). "Response to comment on Hexapod origins: monophyletic or paraphyletic?" (PDF). Science. 301 (5639): 1482. doi:10.1126/science.1087632. S2CID 82407120.
- Gao, Yan; Bu, Yun & Luan, Yun-Xia (2008). "Phylogenetic relationships of basal hexapods reconstructed from nearly complete 18S and 28S rRNA gene sequences" (PDF). Zoological Science. 25 (11): 1139–1145. doi:10.2108/zsj.25.1139. PMID 19267625. S2CID 10783597.
- Hassanin, Alexandre (2006). "Phylogeny of Arthropoda inferred from mitochondrial sequences: strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution" (PDF). Molecular Phylogenetics and Evolution. 38 (1): 100–116. doi:10.1016/j.ympev.2005.09.012. PMID 16290034.
- Davies, W. Maldwyn (1927). "On the tracheal system of Collembola, with special reference to that of Sminthurus viridis, Lubb" (PDF). Quarterly Journal of Microscopical Science. 71 (281): 15–30.
- Hopkin, Stephen P. (1997). Biology of the springtails (Insecta: Collembola). Oxford: Oxford University Press.
- Piper, Ross (2007). Extraordinary animals: an encyclopedia of curious and unusual animals. Santa Barbara, California: Greenwood Press. ISBN 9780313339226. OCLC 124074839.
- "The incredible shrinking springtail". Science. 341 (6149): 945. 30 August 2013. doi:10.1126/science.341.6149.945-a.
- Multicellular Animals: Volume II: The Phylogenetic System of the Metazoa
- Coleman, David C.; Wall, Diana H. (2015-01-01), Paul, Eldor A. (ed.), "Chapter 5 - Soil Fauna: Occurrence, Biodiversity, and Roles in Ecosystem Function", Soil Microbiology, Ecology and Biochemistry (Fourth Edition), Academic Press, pp. 111–149, ISBN 978-0-12-415955-6, retrieved 2020-03-16
- Potapov, Anton A.; Semenina, Eugenia E.; Korotkevich, Anastasiya Yu.; Kuznetsova, Natalia A.; Tiunov, Alexei V. (2016-10-01). "Connecting taxonomy and ecology: Trophic niches of collembolans as related to taxonomic identity and life forms". Soil Biology and Biochemistry. 101: 20–31. doi:10.1016/j.soilbio.2016.07.002. ISSN 0038-0717.
- Jordana, Rafael; Baquero Martín, Enrique; Ledesma, Enrique; Sendra, Alberto; Ortuño, Vicente (2020-02-01). "Poduromorpha (Collembola) from a sampling in the mesovoid shallow substratum of the Sierra de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and Biogeography". Zoologischer Anzeiger. 285: 81–96. doi:10.1016/j.jcz.2020.02.001. S2CID 214349457.
- Xiong, Y; Gao, Y; Yin, W; Luan, Y (2008). "Molecular phylogeny of Collembola inferred from ribosomal RNA genes". Molecular Phylogenetics and Evolution. 49 (3): 728–735. doi:10.1016/j.ympev.2008.09.007. ISSN 1055-7903. PMID 18835455.
- Leo, Chiara; Carapelli, Antonio; Cicconardi, Francesco; Frati, Francesco; Nardi, Francesco (2019). "Mitochondrial Genome Diversity in Collembola: Phylogeny, Dating and Gene Order". Diversity. 11 (9): 169. doi:10.3390/d11090169. ISSN 1424-2818.
- Sun, Xin; Ding, Yinhuan; Orr, Michael C.; Zhang, Feng (2020). "Streamlining universal single‐copy orthologue and ultraconserved element design: A case study in Collembola". Molecular Ecology Resources. 20 (3): 706–717. doi:10.1111/1755-0998.13146. ISSN 1755-098X. PMID 32065730. S2CID 211133755.
- "Checklist of the Collembola". Retrieved January 2, 2016.
- Sánchez-García, Alba & Engel, Michael S. (2016). "Long-term stasis in a diverse fauna of Early Cretaceous springtails (Collembola: Symphypleona)" (PDF). Journal of Systematic Palaeontology: 1–25.
- Daly, Howell V.; Doyen, John T. & Purcell, Alexander H. (1998). Introduction to insect biology and diversity (2nd ed.). New York: Oxford University Press. ISBN 978-0-19-510033-4. OCLC 925231875.
- "Hexapoda. Insects, springtails, diplurans, and proturans". Tree of Life Web Project. January 1, 2002. Retrieved March 26, 2017.
- Mari Mutt, José A. (1983). "Collembola in amber from the Dominican Republic" (PDF). Proceedings of the Entomological Society of Washington. 85 (3): 575–587.
- Nel, André; De Ploëg, Gaėl; Milliet, Jacqueline; Menier, Jean-Jacques & Waller, Alain (2004). "The French ambers: a general conspectus and the Lowermost Eocene amber deposit of Le Quesnoy in the Paris Basin" (PDF). Geologica Acta. 2 (1): 3–8.
- Penney, David; McNeil, Andrew; Green, David I.; Bradley, Robert S.; Jepson, James E.; Withers, Philip J. & Preziosi, Richard F. (2012). "Ancient Ephemeroptera-Collembola symbiosis fossilized in amber predicts contemporary phoretic associations". PLOS ONE. 7 (10): e47651. Bibcode:2012PLoSO...747651P. doi:10.1371/journal.pone.0047651. PMC 3474712. PMID 23082186.
- Christiansen, Kenneth & Nascimbene, Paul (2006). "Collembola (Arthropoda, Hexapoda) from the mid Cretaceous of Myanmar (Burma)" (PDF). Cretaceous Research. 27 (3): 318–33. doi:10.1016/j.cretres.2005.07.003.
- Koehler, Philip G.; Aparicio, M. L. & Pfiester, Margaret (July 2011). "Springtails" (PDF). Gainesville, Florida: University of Florida IFAS Extension. Retrieved March 26, 2017.
- Sechi, Valentina; D'Annibale, Alessandra; Ambus, Per; Sárossy, Zsuzsa; Krogh, Paul Henning; Eriksen, Jørgen; Holmstrup, Martin (2014-07-01). "Collembola feeding habits and niche specialization in agricultural grasslands of different composition". Soil Biology and Biochemistry. 74: 31–38. doi:10.1016/j.soilbio.2014.02.019. ISSN 0038-0717.
- Rusek, Josef (September 1998). "Biodiversity of Collembola and their functional role in the ecosystem". Biodiversity and Conservation. 7 (9): 1207–1219. doi:10.1023/a:1008887817883. ISSN 0960-3115. S2CID 22883809.
- Chen, Benrong; Snider, Richard J. & Snider, Renate M. (1996). "Food consumption by Collembola from northern Michigan deciduous forest" (PDF). Pedobiologia. 40 (2): 149–161.
- Koehler, H.H. (1999), "Predatory mites (Gamasina, Mesostigmata)", Invertebrate Biodiversity as Bioindicators of Sustainable Landscapes, Elsevier, pp. 395–410, doi:10.1016/b978-0-444-50019-9.50022-4, ISBN 978-0-444-50019-9, retrieved 2022-10-06
- City of Austin and Travis County (September 3, 2020). "Support for Maintaining Endangered Status For the Bone Cave Harvestman (Texella reyesi)". Retrieved October 6, 2022.
- Hopkin, Stephen P. (1997). "The biology of the Collembola (springtails): the most abundant insects in the world" (PDF). Natural History Museum. Retrieved January 2, 2016.
- Ponge, Jean-François (1991). "Food resources and diets of soil animals in a small area of Scots pine litter" (PDF). Geoderma. 49 (1–2): 33–62. Bibcode:1991Geode..49...33P. CiteSeerX 10.1.1.635.8529. doi:10.1016/0016-7061(91)90090-G.
- Hopkin, Stephen P. (1997). Biology of the Springtails: (Insecta: Collembola). OUP Oxford. p. 127. ISBN 978-0-19-158925-6.
- Ponge, Jean-François; Arpin, Pierre; Sondag, Francis & Delecour, Ferdinand (1997). "Soil fauna and site assessment in beech stands of the Belgian Ardennes" (PDF). Canadian Journal of Forest Research. 27 (12): 2053–2064. doi:10.1139/cjfr-27-12-2053.
- Ponge, Jean-François (1993). "Biocenoses of Collembola in atlantic temperate grass-woodland ecosystems" (PDF). Pedobiologia. 37 (4): 223–244.
- Lyford, Walter H. (1975). "Overland migration of Collembola (Hypogastrura nivicola Fitch) colonies" (PDF). American Midland Naturalist. 94 (1): 205–209. doi:10.2307/2424550. JSTOR 2424550.
- Shaw, Peter; Ozanne, Claire; Speight, Martin & Palmer, Imogen (2007). "Edge effects and arboreal Collembola in coniferous plantations" (PDF). Pedobiologia. 51 (4): 287–293. doi:10.1016/j.pedobi.2007.04.010.
- Zettel, Jürg; Zettel, Ursula & Egger, Beatrice (2000). "Jumping technique and climbing behaviour of the collembolan Ceratophysella sigillata (Collembola: Hypogastruridae)" (PDF). European Journal of Entomology. 97 (1): 41–45. doi:10.14411/eje.2000.010.
- Didden, Wim A. M. (1987). "Reactions of Onychiurus fimatus (Collembola) to loose and compact soil: methods and first results" (PDF). Pedobiologia. 30 (2): 93–100. doi:10.1016/S0031-4056(23)00360-8. S2CID 258140448.
- Rodgers, Denis J. & Kitching, Rodger L. (1998). "Vertical stratification of rainforest collembolan (Collembola: Insecta) assemblages: description of ecological patterns and hypotheses concerning their generation". Ecography. 21 (4): 392–400. CiteSeerX 10.1.1.476.6663. doi:10.1111/j.1600-0587.1998.tb00404.x. S2CID 85133291.
- Bowden, John; Haines, Ian H. & Mercer, D. (1976). "Climbing Collembola". Pedobiologia. 16 (4): 298–312. doi:10.1016/S0031-4056(23)02178-9. S2CID 258572651.
- Krab, Eveline J.; Oorsprong, Hilde; Berg, Matty P. & Cornelissen, Johannes H. C. (2010). "Turning northern peatlands upside down: disentangling microclimate and substrate quality effects on vertical distribution of Collembola" (PDF). Functional Ecology. 24 (6): 1362–1369. doi:10.1111/j.1365-2435.2010.01754.x.
- Nickerl, Julia; Helbig, Ralf; Schulz, Hans-Jürgen; Werner, Carsten & Neinhuis, Christoph (2013). "Diversity and potential correlations to the function of Collembola cuticle structures" (PDF). Zoomorphology. 132 (2): 183–195. doi:10.1007/s00435-012-0181-0. S2CID 14442664.
- Holmstrup, Martin & Bayley, Mark (2013). "Protaphorura tricampata, a euedaphic and highly permeable springtail that can sustain activity by osmoregulation during extreme drought". Journal of Insect Physiology. 59 (11): 1104–1110. doi:10.1016/j.jinsphys.2013.08.015. PMID 24035747.
- Verhoef, Herman A. (1984). "Releaser and primer pheromones in Collembola" (PDF). Journal of Insect Physiology. 30 (8): 665–670. doi:10.1016/0022-1910(84)90052-0.
- Benoit, Joshua B.; Elnitsky, Michael A.; Schulte, Glen G.; Lee, Richard E. Jr & Denlinger, David L. (2009). "Antarctic Collembolans use chemical signals to promote aggregation and egg laying" (PDF). Journal of Insect Behavior. 22 (2): 121–133. doi:10.1007/s10905-008-9159-7. S2CID 914560.
- Prinzing, Andreas; D'Haese, Cyrille A.; Pavoine, Sandrine & Ponge, Jean-François (2014). "Species living in harsh environments have low clade rank and are localized on former Laurasian continents: a case study of Willemia (Collembola)" (PDF). Journal of Biogeography. 41 (2): 353–365. doi:10.1111/jbi.12188. S2CID 86619537.
- Verhoef, Herman A. (1981). "Water balance in Collembola and its relation to habitat selection: water content, haemolymph osmotic pressure and transpiration during an instar". Journal of Insect Physiology. 27 (11): 755–760. doi:10.1016/0022-1910(81)90065-2.
- Leinaas, Hans Petter (1983). "Synchronized moulting controlled by communication in group-living Collembola". Science. 219 (4581): 193–195. Bibcode:1983Sci...219..193P. doi:10.1126/science.219.4581.193. PMID 17841689. S2CID 41604935.
- Wilson, Jane M. (1982). "A review of world Troglopedetini (Insecta, Collembola, Paronellidae), including an identification table and descriptions of new species" (PDF). Cave Science: Transactions of the British Cave Research Association. 9 (3): 210–226.
- Palacios-Vargas, José G. & Wilson, Jane (1990). "Troglobius coprophagus, a new genus and species of cave collembolan from Madagascar with notes on its ecology" (PDF). International Journal of Speleology. 19 (1–4): 67–73. doi:10.5038/1827-806x.19.1.6.
- Jordana, Rafael; Baquero, Enrique; Reboleira, Sofía & Sendra, Alberto (2012). "Reviews of the genera Schaefferia Absolon, 1900, Deuteraphorura Absolon, 1901, Plutomurus Yosii, 1956 and the Anurida Laboulbène, 1865 species group without eyes, with the description of four new species of cave springtails (Collembola) from Krubera-Voronya cave, Arabika Massif, Abkhazia" (PDF). Terrestrial Arthropod Reviews. 5 (1): 35–85. doi:10.1163/187498312X622430. hdl:10171/27607.
- Salmon, Sandrine; Ponge, Jean-François & Van Straalen, Nico (2002). "Ionic identity of pore water influences pH preference in Collembola" (PDF). Soil Biology and Biochemistry. 34 (11): 1663–1667. doi:10.1016/S0038-0717(02)00150-5.
- Loranger, Gladys; Bandyopadhyaya, Ipsa; Razaka, Barbara & Ponge, Jean-François (2001). "Does soil acidity explain altitudinal sequences in collembolan communities?" (PDF). Soil Biology and Biochemistry. 33 (3): 381–393. doi:10.1016/S0038-0717(00)00153-X. S2CID 84523833.
- Faber, Jack H. & Joosse, Els N.G. (1993). "Vertical distribution of Collembola in a Pinus nigra organic soil". Pedobiologia. 37 (6): 336–350.
- Detsis, Vassilis (2000). "Vertical distribution of Collembola in deciduous forests under Mediterranean climatic conditions" (PDF). Belgian Journal of Zoology. 130 (Supplement 1): 57–61.
- "Isotomurus palustris (Muller, 1776)". Retrieved April 2, 2017.
- Pichard, Sylvain (1973). "Contribution à l'étude de la biologie de Podura aquatica (Linné) Collembole". Bulletin Biologique de la France et de la Belgique (in French). 107 (4): 291–299.
- Ponge, Jean-François; Gillet, Servane; Dubs, Florence; Fédoroff, Éric; Haese, Lucienne; Sousa, José Paulo & Lavelle, Patrick (2003). "Collembolan communities as bioindicators of land use intensification" (PDF). Soil Biology and Biochemistry. 35 (6): 813–826. doi:10.1016/S0038-0717(03)00108-1. hdl:10316/3875. S2CID 86289373.
- Ponge, Jean-François; Dubs, Florence; Gillet, Servane; Sousa, José Paulo & Lavelle, Patrick (2006). "Decreased biodiversity in soil springtail communities: the importance of dispersal and landuse history in heterogeneous landscapes" (PDF). Soil Biology and Biochemistry. 38 (5): 1158–1161. doi:10.1016/j.soilbio.2005.09.004.
- Cristofoli, Sara & Mahy, Grégory (2010). "Colonisation credit in recent wet heathland butterfly communities". Insect Conservation and Diversity. 3 (2): 83–91. doi:10.1111/j.1752-4598.2009.00075.x. S2CID 86558143.
- Heiniger, Charlène; Barot, Sébastien; Ponge, Jean-François; Salmon, Sandrine; Botton-Divet, Léo; Carmignac, David & Dubs, Florence (2014). "Effect of habitat spatiotemporal structure on collembolan diversity" (PDF). Pedobiologia. 57 (2): 103–117. doi:10.1016/j.pedobi.2014.01.006.
- Shaw, Michael W. & Haughs, G. M. (1983). "Damage to potato foliage by Sminthurus viridis (L.)" (PDF). Plant Pathology. 32 (4): 465–466. doi:10.1111/j.1365-3059.1983.tb02864.x.
- Bishop, Alan L.; Harris, Anne M. & McKenzie, Harry J. (2001). "Distribution and ecology of the lucerne flea, Sminthurus viridis (L.) (Collembola: Sminthuridae), in irrigated lucerne in the Hunter dairying region of New South Wales" (PDF). Australian Journal of Entomology. 40 (1): 49–55. doi:10.1046/j.1440-6055.2001.00202.x.
- "Lucerne Flea". State of Victoria. 2017. Retrieved June 2, 2018.
- Baker, A. N. & Dunning, Andrew R. (1975). "Association of populations of onychiurid Collembola with damage to sugar-beet seedlings" (PDF). Plant Pathology. 24 (3): 150–154. doi:10.1111/j.1365-3059.1975.tb01882.x.
- Klironomos, John N. & Moutoglis, Peter (1999). "Colonization of nonmycorrhizal plants by mycorrhizal neighbours as influenced by the collembolan, Folsomia candida" (PDF). Biology and Fertility of Soils. 29 (3): 277–281. doi:10.1007/s003740050553. S2CID 11014525.
- Sabatini, Maria Agnese & Innocenti, Gloria (2001). "Effects of Collembola on plant-pathogenic fungus interactions in simple experimental systems" (PDF). Biology and Fertility of Soils. 33 (1): 62–66. doi:10.1007/s003740000290. S2CID 9273050.
- Shiraishi, Hiroyoshi; Enami, Yoshinari & Okano, Seigo (2003). "Folsomia hidakana (Collembola) prevents damping-off disease in cabbage and Chinese cabbage by Rhizoctonia solani" (PDF). Pedobiologia. 47 (1): 33–38. doi:10.1078/0031-4056-00167.
- Ponge, Jean-François & Charpentié, Marie-José (1981). "Étude des relations microflore-microfaune: expériences sur Pseudosinella alba (Packard), Collembole mycophage" (PDF). Revue d'Écologie et de Biologie du Sol (in French). 18: 291–303.
- Lartey, Robert T.; Curl, Elroy A.; Peterson, Curt M. & Harper, James D. (1989). "Mycophagous grazing and food preference of Proisotoma minuta (Collembola: Isotomidae) and Onychiurus encarpatus (Collembola: Onychiuridae)". Environmental Entomology. 18 (2): 334–337. doi:10.1093/ee/18.2.334.
- Janssens, Frans & Christiansen, Kenneth A. (November 22, 2007). "Synanthropic Collembola, springtails in association with Man". Checklist of the Collembola. Retrieved April 9, 2017.
- Berenbaum, May (2005). "Face time" (PDF). American Entomologist. 51 (2): 68–69. doi:10.1093/ae/51.2.68.
- Christiansen, Kenneth & Bernard, Ernest C. (2008). "Critique of the article "Collembola (Springtails) (Arthropoda: Hexapoda: Entognatha) found in scrapings from individuals diagnosed with delusory parasitosis"". Entomological News. 119 (5): 537–540. doi:10.3157/0013-872x-119.5.537. S2CID 83870937.
- Lim, Christopher Seng-Hong; Lim, Scott L.; Chew, Fook Tim & Deharveng, Louis (2009). "Collembola are unlikely to cause human dermatitis". Journal of Insect Science. 9 (3): 1–5. doi:10.1673/031.009.0301. PMC 3011850. PMID 19611235.
- Shelomi, Matan (2013). "Evidence of photo manipulation in a delusional parasitosis paper" (PDF). Journal of Parasitology. 99 (3): 583–585. doi:10.1645/12-12.1. PMID 23198757. S2CID 6473251. Retrieved 29 January 2019.
- "Insects as Biological Weapons - Insects, Disease, and History | Montana State University". www.montana.edu. Retrieved 2022-10-06.
- Lockwood, Jeffrey A. (2009). Six-legged soldiers: using insects as weapons of war (1st ed.). Oxford, UK: Oxford University Press. ISBN 978-0-19-533305-3.
- "The Top 5 Bioactive Clean Up Crew". Northampton Reptile Centre. 16 December 2015. Retrieved 6 July 2021.
- Fountain, Michelle T. & Hopkin, Steve P. (2001). "Continuous monitoring of Folsomia candida (Insecta: Collembola) in a metal exposure test" (PDF). Ecotoxicology and Environmental Safety. 48 (3): 275–286. doi:10.1006/eesa.2000.2007. PMID 11222037.
- ISO 11267 (2014). "Soil quality. Inhibition of reproduction of Collembola (Folsomia candida) by soil contaminants". Geneva: International Organization for Standardization.
- Krogh, Paul Henning (August 1, 2008). "Toxicity testing with the collembolans Folsomia fimetaria and Folsomia candida and the results of a ringtest" (PDF). Retrieved April 17, 2017.
- Lors, Christine; Martínez Aldaya, Maite; Salmon, Sandrine & Ponge, Jean-François (2006). "Use of an avoidance test for the assessment of microbial degradation of PAHs" (PDF). Soil Biology and Biochemistry. 38 (8): 2199–2204. doi:10.1016/j.soilbio.2006.01.026.
- ISO 17512-2 (2011). "Soil quality. Avoidance test for determining the quality of soils and effects of chemicals on behaviour. Part 2: Test with collembolans (Folsomia candida)". Geneva: International Organization for Standardization.
- Chauvat, Matthieu & Ponge, Jean-François (2002). "Colonization of heavy metal-polluted soils by collembola: preliminary experiments in compartmented boxes" (PDF). Applied Soil Ecology. 21 (2): 91–106. doi:10.1016/S0929-1393(02)00087-2.
- Ponge, Jean-François; Bandyopadhyaya, Ipsa & Marchetti, Valérie (2002). "Interaction between humus form and herbicide toxicity to Collembola (Hexapoda)" (PDF). Applied Soil Ecology. 20 (3): 239–253. doi:10.1016/S0929-1393(02)00026-4.
- Nota, Benjamin; Timmermans, Martijn J.T.N.; Franken, Oscar; Montagne-Wajer, Kora; Mariën, Janine; De Boer, Muriel E.; De Boer, Tjalf E.; Ylstra, Bauke; Van Straalen, Nico M. & Roelofs, Dick (2008). "Gene expression analysis of Collembola in cadmium containing soil" (PDF). Environmental Science and Technology. 42 (21): 8152–8157. Bibcode:2008EnST...42.8152N. doi:10.1021/es801472r. PMID 19031917.
- Nota, Benjamin; Bosse, Mirte; Ylstra, Bauke; Van Straalen, Nico M. & Roelofs, Dick (2009). "Transcriptomics reveals extensive inducible biotransformation in the soil-dwelling invertebrate Folsomia candida exposed to phenanthrene". BMC Genomics. 10: 236. doi:10.1186/1471-2164-10-236. PMC 2688526. PMID 19457238.
- Kim, Shin Woong & An, Youn-Joo (2014). "Jumping behavior of the springtail Folsomia candida as a novel soil quality indicator in metal-contaminated soils". Ecological Indicators. 38: 67–71. doi:10.1016/j.ecolind.2013.10.033.
- Nielsen, Uffe N. & Wall, Diana H. (2013). "The future of soil invertebrate communities in polar regions: different climate change responses in the Arctic and Antarctic?". Ecology Letters. 16 (3): 409–419. doi:10.1111/ele.12058. PMID 23278945.
- Coulson, Stephen James; Hodkinson, Ian D.; Woolley, Christopher; Webb, Nigel R.; Block, William; Worland, M. Rodger; Bale, Jeff S. & Strathdee, Andrew T. (1996). "Effects of experimental temperature elevation on high-arctic soil microarthropod populations" (PDF). Polar Biology. 16 (2): 147–153. doi:10.1007/BF02390435. S2CID 13826340.
- Sjursen, Heidi; Michelsen, Anders & Jonasson, Sven (2005). "Effects of long-term soil warming and fertilisation on microarthropod abundances in three sub-arctic ecosystems" (PDF). Applied Soil Ecology. 30 (3): 148–161. doi:10.1016/j.apsoil.2005.02.013.
- Dollery, Rebecca; Hodkinson, Ian D. & Jónsdóttir, Ingibjörg S. (2006). "Impact of warming and timing of snow melt on soil microarthropod assemblages associated with Dryas-dominated plant communities on Svalbard" (PDF). Ecography. 29 (1): 111–119. doi:10.1111/j.2006.0906-7590.04366.x.
- Hågvar, Sigmund & Klanderud, Kari (2009). "Effect of simulated environmental change on alpine soil arthropods" (PDF). Global Change Biology. 15 (12): 2972–2980. Bibcode:2009GCBio..15.2972H. CiteSeerX 10.1.1.658.1606. doi:10.1111/j.1365-2486.2009.01926.x. S2CID 36287733.
- Alatalo, Juha M.; Jägerbrand, Annika K. & Čuchta, Peter (2015). "Collembola in three alpine subarctic sites resistant to twenty years of experimental warming". Scientific Reports. 5 (18161): 18161. Bibcode:2015NatSR...518161A. doi:10.1038/srep18161. PMC 4680968. PMID 26670681.
- Kardol, Paul; Reynolds, W. Nicholas; Norby, Richard J. & Classen, Aimée T. (2011). "Climate change effects on soil microarthropod abundance and community structure" (PDF). Applied Soil Ecology. 47 (1): 37–44. doi:10.1016/j.apsoil.2010.11.001.
- Harte, John; Rawa, Agnieszka & Price, Vanessa (1996). "Effects of manipulated soil microclimate on mesofaunal biomass and diversity" (PDF). Soil Biology and Biochemistry. 28 (3): 313–322. doi:10.1016/0038-0717(95)00139-5.
- Waldorf, Elizabeth S. (1974). "Sex pheromone in the springtail Sinella curviseta". Environmental Entomology. 3 (6): 916–918. doi:10.1093/ee/3.6.916.
- Kozlowski, Marek Wojciech & Aoxiang, Shi (2006). "Ritual behaviors associated with spermatophore transfer in Deuterosminthurus bicinctus (Collembola : Bourletiellidae)". Journal of Ethology. 24 (2): 103–110. doi:10.1007/s10164-005-0162-6. S2CID 12056710.
- Simon, Jean-Christophe; Delmote, François; Rispe, Claude & Crease, Teresa (2003). "Phylogenetic relationships between parthenogens and their sexual relatives: the possible routes to parthenogenesis in animals" (PDF). Biological Journal of the Linnean Society. 79 (1): 151–153. doi:10.1046/j.1095-8312.2003.00175.x.
- Czarnetzki, Alice B. & Tebbe, Christoph C. (2004). "Detection and phylogenetic analysis of Wolbachia in Collembola" (PDF). Environmental Microbiology. 6 (1): 35–44. doi:10.1046/j.1462-2920.2003.00537.x. PMID 14686939.
- Werren, John H.; Zhang, Wan & Guo, Li Rong (1995). "Evolution and phylogeny of Wolbachia: reproductive parasites of arthropods" (PDF). Proceedings of the Royal Society B. 261 (1360): 55–63. Bibcode:1995RSPSB.261...55W. doi:10.1098/rspb.1995.0117. JSTOR 50047. PMID 7644549. S2CID 8585323.
- Fenn, Katelyn & Blaxter, Mark (2004). "Are filarial nematode Wolbachia obligate mutualist symbionts?" (PDF). Trends in Ecology and Evolution. 19 (4): 163–166. doi:10.1016/j.tree.2004.01.002. PMID 16701248.
External links
- . Encyclopædia Britannica. Vol. 25 (11th ed.). 1911.
- Checklist of the Collembola of the World
- Summary information about the distribution and ecology of Collembola (springtails) in the UK and Ireland
- General information on Collembola
- General information on Collembola
- A small lecture from Steve Hopkin
- General information on Collembola, with many macrophotographs from Dutch springtails
- Collembola species currently recorded from South Africa