Cysteine-rich protein
Cysteine-rich proteins (also cysteine-rich peptide, CRP, disulphide-rich peptide) are small proteins that contain a large number of cysteines. These cysteines either cross-link to form disulphide bonds, or bind metal ions by chelation, stabilising the protein's tertiary structure.[1][2][3] CRPs include a highly conserved secretion peptide signal at the N-terminus and a cysteine-rich region at the C-terminus.[4]
Structure
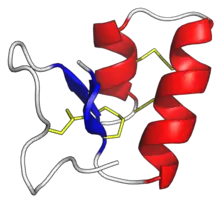
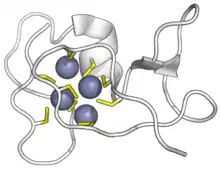
Disulphides
In an oxidising environment cysteines cross-link to form disulphide bonds. CRPs that form these typically have an even number of cysteines.[5]
Functions in plants
CRPs are numerous in plants, with 756 CRP-encoding genes in the Arabidopsis thaliana genome.[7] Several CRPs bind known receptors,[8] but most CRP signaling mechanisms and protein interactions are uncharacterized. Characterized CRPs function as short-range intercellular signals during processes such as plant defense, bacterial symbiosis, stomatal patterning, fertilization, vegetative tissue development, and seed development.[4]
Many CRPs function in plant defense. Defensins, a major class of CRP with an eight-cysteine motif forming four disulfide bridges,[9] are involved in pathogen response.[4] Other putative antimicrobial CRPs include lipid transfer proteins, thionins, knottins, heveins, and snakins. Additionally, some CRPs have allergenic, ɑ-amylase inhibitory, or protease inhibitory functions that deter herbivores.[9]
In plant reproduction, CRPs are involved in pollen tube growth and guidance[10] and early embryo patterning,[11] in addition to other functions. Among those involved in pollen tube attraction are the LUREs, a group of ovular pollen-tube attractants in Arabidopsis thaliana and Torenia fournieri[12] that preferentially attract conspecific pollen,[10] and STIG1, a CRP expressed in the stigma of Solanum lycopersicum that interacts with the pollen-specific receptor PRK2.[8] In early embryo development, CRPs such as ESF1 are necessary for suspensor development and normal seed morphology.[11]
References
- Cheek S, Krishna SS, Grishin NV (May 2006). "Structural classification of small, disulfide-rich protein domains". Journal of Molecular Biology. 359 (1): 215–37. doi:10.1016/j.jmb.2006.03.017. PMID 16618491.
- Arolas JL, Aviles FX, Chang JY, Ventura S (May 2006). "Folding of small disulfide-rich proteins: clarifying the puzzle". Trends in Biochemical Sciences. 31 (5): 292–301. doi:10.1016/j.tibs.2006.03.005. PMID 16600598. S2CID 30709875.
- Metallothioneins and related chelators. Sigel, Astrid., Sigel, Helmut., Sigel, Roland K. O. Cambridge: Royal Society of Chemistry. 2009. ISBN 978-1-84755-953-1. OCLC 429670531.
{{cite book}}: CS1 maint: others (link) - Marshall E, Costa LM, Gutierrez-Marcos J (March 2011). "Cysteine-rich peptides (CRPs) mediate diverse aspects of cell-cell communication in plant reproduction and development". Journal of Experimental Botany. 62 (5): 1677–86. doi:10.1093/jxb/err002. PMID 21317212.
- Lavergne, Vincent; J. Taft, Ryan; F. Alewood, Paul (2012-08-01). "Cysteine-Rich Mini-Proteins in Human Biology". Current Topics in Medicinal Chemistry. 12 (14): 1514–1533. doi:10.2174/156802612802652411. ISSN 1568-0266. PMID 22827521.
- Maret, Wolfgang (2008-05-01). "Metallothionein redox biology in the cytoprotective and cytotoxic functions of zinc". Experimental Gerontology. Zinc and Ageing (ZINCAGE Project). 43 (5): 363–369. doi:10.1016/j.exger.2007.11.005. ISSN 0531-5565. PMID 18171607. S2CID 19456564.
- Huang Q, Dresselhaus T, Gu H, Qu LJ (June 2015). "Active role of small peptides in Arabidopsis reproduction: Expression evidence". Journal of Integrative Plant Biology. 57 (6): 518–21. doi:10.1111/jipb.12356. PMID 25828584.
- Huang WJ, Liu HK, McCormick S, Tang WH (June 2014). "Tomato Pistil Factor STIG1 Promotes in Vivo Pollen Tube Growth by Binding to Phosphatidylinositol 3-Phosphate and the Extracellular Domain of the Pollen Receptor Kinase LePRK2". The Plant Cell. 26 (6): 2505–2523. doi:10.1105/tpc.114.123281. PMC 4114948. PMID 24938288.
- Silverstein KA, Moskal WA, Wu HC, Underwood BA, Graham MA, Town CD, VandenBosch KA (July 2007). "Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants". The Plant Journal. 51 (2): 262–80. doi:10.1111/j.1365-313X.2007.03136.x. PMID 17565583.
- Zhong S, Liu M, Wang Z, Huang Q, Hou S, Xu YC, et al. (May 2019). "Arabidopsis". Science. 364 (6443): eaau9564. doi:10.1126/science.aau9564. PMC 7184628. PMID 31147494.
- Costa LM, Marshall E, Tesfaye M, Silverstein KA, Mori M, Umetsu Y, et al. (April 2014). "Central cell-derived peptides regulate early embryo patterning in flowering plants". Science. 344 (6180): 168–72. Bibcode:2014Sci...344..168C. doi:10.1126/science.1243005. PMID 24723605. S2CID 3234638.
- Kanaoka MM, Higashiyama T (December 2015). "Peptide signaling in pollen tube guidance". Current Opinion in Plant Biology. 28: 127–36. doi:10.1016/j.pbi.2015.10.006. PMID 26580200.