DCL2
DCL2 (an abbreviation of Dicer-like 2) is a gene in plants that codes for the DCL2 protein, a ribonuclease III enzyme involved in processing exogenous double-stranded RNA (dsRNA) into 22 nucleotide small interference RNAs (siRNAs).[1]
| Endoribonuclease Dicer homolog 2 | |||||||
|---|---|---|---|---|---|---|---|
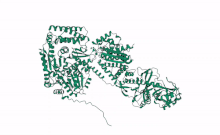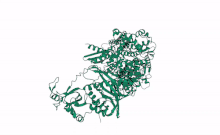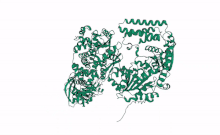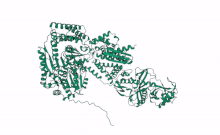 Cartoon representation of Arabidopsis DCL2, Based on computational predictions using Alphafold2 and rendered with open software Mol Star * (https://alphafold.ebi.ac.uk/entry/Q3EBC8, https://molstar.org/viewer/) | |||||||
| Identifiers | |||||||
| Organism | |||||||
| Symbol | DCL2 | ||||||
| Alt. symbols | AT3G03300 | ||||||
| UniProt | Q3EBC8 | ||||||
| |||||||
Diverse sources of dsRNAs have been characterized, broadly classified as exogenous or endogenous. A classical example of exogenous derived dsRNAs are the viral genomes release during infection, specially from those double-stranded RNA viruses, where the cleavage of dsRNA produce small RNA products called viral siRNAs or vsi-RNAs.[2] Other examples of exogenous source of dsRNAs are transgenic with several insertion loci along the plant hos genome.[3] DCL2 also process endogenous sources as double-stranded RNAs derived of cis-natural antisense transcripts, generating 22nt short interfering RNA (natsi-RNAs); however, the biological relevance, evolutionary conservation, and experimental validation of natsi-RNAs remains controversial.[4]
Function
Dicer proteins belongs to the RNaseIII-like family, a gene family with highly conserved endonuclease in eukaryotes, with procaryotes representatives.[5] In Arabidopsis and most of land Plants, there are mainly four Dicer-like proteins (DCL): DCL1, DCL2, DCL3, and DCL4. They all contain five domains, following the order from N-terminus to C-terminus: DEXD-helicase, helicase-C, domain of unknown function 283 (DUF283), Piwi/Argonaute/Zwille (PAZ) domain, two tandem RNase III domains, and one or two dsRNA-binding domains (dsRBDs).[5] In general, the helicase domain of dicer-like proteins utilizes ATP hydrolysis to facilitate the unwinding of dsRNA.[5] The DUF283 domain have been recently associated as a protein domain involve in facilitation of RNA-RNA base pairing and RNA-binding.[6] The PAZ and RNase III domains are essential for dsRNA cleavage via the recognition of dsRNA ends by PAZ domain, the RNase III domains cuts one of the strands of dsRNA.[7]
DCL2 plays an essential role in transitive silencing of transgenes by processing secondary siRNAs, including trans-acting siRNA.[4] To do so, it does requires DCL4 and RDR6, which amplifies the silencing by using the mRNA target of the DCL2's generated 22nt siRNA, as substrate to generate secondary siRNAs, providing an efficient mechanism for long-distance silencing, in a phenomenon called transitivity of RNA silencing.[8]
DCL2 may participate as well with DCL3 in the production of 24 nucleotide repeat-associated siRNAs (ra-siRNAs) derived from heterochromatic regions, genomic regions silenced by the presence of DNA repetitive elements such as transposons.[9]
Transitive and systemic RNA silencing
A key difference between DCL1 and others DCLs 2,3 and 4 proteins is the amplification capacity of the pathways specific for the later protein. The involvement of RDR proteins extends the small RNA-target complex beyond the original trigger-spot. The subset of siRNA used in signal amplification are called transitive or secondary siRNAs and the process of amplification is called transitivity.[8] The amplification propagates the secondary siRNA and its target specific silencing activity from one tissue to another, eventually reaching the whole plant's tissues, in a process called systemic silencing.[8]
References
- Jin L, Chen M, Xiang M, Guo Z (February 2022). "RNAi-Based Antiviral Innate Immunity in Plants". Viruses. 14 (2): 432. doi:10.3390/v14020432. PMC 8875485. PMID 35216025.
- Sanan-Mishra N, Abdul Kader Jailani A, Mandal B, Mukherjee SK (2021-03-02). "Secondary siRNAs in Plants: Biosynthesis, Various Functions, and Applications in Virology". Frontiers in Plant Science. 12: 610283. doi:10.3389/fpls.2021.610283. PMC 7960677. PMID 33737942.
- Stam M (January 1997). "Review Article: The Silence of Genes in Transgenic Plants". Annals of Botany. 79 (1): 3–12. doi:10.1006/anbo.1996.0295.
- Meyers BC, Zhan J, Shevela D, Slotkin KR (2022), Plant Small RNAs: Biogenesis and Functions, Agrisera Educational Poster Collection, Poster 6, 2022, Figshare, doi:10.6084/m9.figshare.21186073, retrieved 2022-11-06
- Cenik ES, Fukunaga R, Lu G, Dutcher R, Wang Y, Tanaka Hall TM, Zamore PD (April 2011). "Phosphate and R2D2 restrict the substrate specificity of Dicer-2, an ATP-driven ribonuclease". Molecular Cell. 42 (2): 172–184. doi:10.1016/j.molcel.2011.03.002. PMC 3115569. PMID 21419681.
- Szczepanska A, Wojnicka M, Kurzynska-Kokorniak A (August 2021). "The Significance of the DUF283 Domain for the Activity of Human Ribonuclease Dicer". International Journal of Molecular Sciences. 22 (16): 8690. doi:10.3390/ijms22168690. PMC 8395393. PMID 34445396.
- Welker NC, Pavelec DM, Nix DA, Duchaine TF, Kennedy S, Bass BL (May 2010). "Dicer's helicase domain is required for accumulation of some, but not all, C. elegans endogenous siRNAs". RNA. 16 (5): 893–903. doi:10.1261/rna.2122010. PMC 2856884. PMID 20354150.
- Choudhary S, Thakur S, Bhardwaj P (August 2019). "Molecular basis of transitivity in plant RNA silencing". Molecular Biology Reports. 46 (4): 4645–4660. doi:10.1007/s11033-019-04866-9. PMID 31098805. S2CID 155103992.
- Benoit M (2020). "Slice and Dice: DCL2 Mediates the Production of 22-Nucleotide siRNAs that Influence Trait Variation in Soybean". The Plant Cell. 32 (12): 3646–3647. doi:10.1105/tpc.20.00884. PMC 7721339. PMID 33093146.