Dinosaur tooth
Dinosaur teeth have been studied since 1822 when Mary Ann Mantell (1795-1869) and her husband Dr Gideon Algernon Mantell (1790-1852) discovered an Iguanodon tooth in Sussex in England. Unlike mammal teeth, individual dinosaur teeth are generally not considered by paleontologists to be diagnostic to the genus or species level for unknown taxa, due morphological convergence and variability between teeth.[1] and many historically named tooth taxa like Paronychodon and Richardoestesia are today considered nomina dubia, and are used as form taxa to refer to isolated teeth from other localities displaced considerably in time and space from the type specimens. However, it is possible to refer isolated teeth to known taxa provided that the tooth morphology is known and the teeth originate from a similar time and place.

Some of the most important anatomical information about dinosaur teeth is collected from polished, microscopically thin sections (histology), including the types of dental tissues present, tooth wear, tooth replacement patterns, how the teeth are attached, and the frequency of replacement. The actual material comprising a dinosaur tooth is not very different to teeth in modern animals. Most significant differences are in how the teeth fit together and continually regrew, with some examples shedding old teeth and others reabsorbing old teeth as they would grind down under chewing throughout a dinosaurs life.
Background

The use of histology in paleontology has traditionally been more focused on examining long bones such as the femur or the humerus. Previous work on long bone histology revealed differences in the growth patterns of polar dinosaurs,[2] identified a case of dwarfism in Europasaurus,[3] reconstructed the life history of Dysalotosaurus by examining multiple specimens of different ontogenetic stages,[4] and suggested that Psittacosaurus underwent a postural change from a quadruped to biped as it matured.[5]
By contrast, dental histology has not been looked at in great detail in dinosaurs until more recently and there has been an increase in interest in this particular sub-field. Histology studies traditionally rely upon the destructive process of creating and examining thin sections under microscopy, often restricting studies to taxa that have plentiful specimens such as isolated teeth or damaged specimens. While non-destructive means of analysis are sometimes possible through the use of scanning electron microscopy (SEM) or micro computed tomography, much anatomical information is difficult to obtain without creating thin sections.[6][7]
Histology
Histological study is microscopic examination, essential to revealing the most important aspects of dinosaur dental anatomy.
Selection
Different specimens will be suitable for looking at particular anatomical features. For example, specimens with teeth intact within the jaws are necessary to study tooth attachment as this information is lost on isolated teeth.[6] On the other hand, isolated teeth would be sufficient if the goal is to examine wear on the tooth surface.
Embedding and sectioning
Thin sections are prepared by first embedding a suitable specimen in epoxy resin. The embedded specimen can then be mounted and cut with a precision saw.[6] The resulting slice is attached to a slide and ground down, then polished, until it is thin enough, with a suitable surface to be examined with a microscope.[6]
Examination
Thin sections are typically examined with a petrographic microscope using plain light and cross-polarized light. Some structures are more easily visible and distinct using one type of light over the other due to differences in their mineral properties. Some specimens can also be examined with a SEM.[7]
Dental anatomy

Various major groups of dinosaurs have been examined through histology, these include the carnivorous theropods and herbivorous groups such as the sauropods, hadrosaurs and ceratopsians.[6][8][9][10][7] Listed below are some of the dental anatomy that has been identified through histology and interpretations on their significance.
Tissue types
There are generally 5 tissue types present in dinosaurs, and these have been found to be identical to those of their closest living non-avian relatives, the crocodilians.[6] One of the most significant findings is that despite differences in their appearance, dinosaur teeth are essentially composed of the same dental tissues found in modern mammals, crocodilians and other amniotes, suggesting that these tissues first evolved in a common ancestor and has been retained ever since.[11][12]
- Enamel - This is the hard coating on the outside of the teeth and typically appears as a clear, thin featureless band on the tooth surface when viewed in cross section.[6] SEM analysis of the surface of dinosaur teeth revealed that their enamel form in prisms similar to mammals and that there is sufficient difference in the enamel microstructure to help pinpoint what group a tooth belonged to, sometimes to the genus level, when only isolated teeth are found.[13] Not all teeth are covered by a prismatic enamel, and in most taxa, prisms are perpendicular to the outer surface of the tooth. Complex arrangements such as visible in mammals are rare.[14][15] Diagenetic alterations modify the structure and composition of both enamel and dentin.[16][17][18][19]
- Dentine - This tissue makes up the bulk of the tooth and is characterized by long thin parallel tubules running throughout the body of the tooth.[6]
- Cementum - This tissue covers the root of a tooth and is an attachment tissue that forms part of the periodontium. It is typically infilled with Sharpey's fibers that help anchor the tooth in place in the socket.[6]
- Periodontal ligament - This is a soft tissue layer between the cementum and the tooth socket. While this is not preserved in fossils, there is always a mineral filled gap that is present in all dinosaur teeth between the cementum and the tooth socket, which infers the presence of soft tissue in life.[6]
- Alveolar bone - This is a type of bone that is typically spongy in appearance and forms the tooth socket itself.[6]
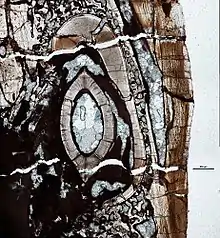
Growth
In some examples viewed in cross section, growth lines can be observed in the dentine of dinosaur teeth. These are known as lines of von Ebner and represent daily deposition of dentine.[20] Counting these lines provides the age of a tooth and comparing the age of the mature tooth to the replacement tooth in a socket provides an estimate of the tooth replacement rate.[20]
The difference in age between the oldest teeth and the youngest teeth is used to determine the rate of tooth replacement.[20]
Tooth replacement pattern
Many dinosaur teeth have been found to have a replacement pattern similar to other reptiles where a replacement tooth grows in the dental lamina on the inside of the jaw before migrating outwards, resorbing part of the growing functional tooth, until ready to erupt and replace it.[6][21]
Tooth attachment
The tooth attachment mode of some dinosaurs has been referred to as thecodonty.[6] This is a condition where the tooth is deeply implanted into the tooth socket with periodontal ligament present, as is the case in crocodilians and mammals.[6][12] In mammals, thecodonty is associated with dental occlusion while in crocodilians it has been proposed as a means to reduce stresses from bite forces.[22][23] Coelophysis possessed neither dental occlusion nor a strong bite, raising questions as to why it possesses thecodonty.[6]
Dental batteries
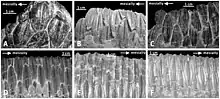
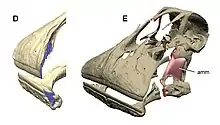
One of the most complex dentition found in dinosaurs are the dental batteries present in hadrosaurs (whose members were dominant species across the planet), Neoceratopsia (for example, Triceratops), and Rebbachisauridae.[24]
These batteries were formed from hundreds of teeth which were stacked in rows upon rows and formed a grinding surface to process plant foods.[24] Histological study of these batteries found that they were not cemented together as previously thought, but that each tooth in the battery was separately moving and supported by ligamenture such that the whole structure was flexible.[9][24] Comparable to shark teeth, dental batteries exhibited polyphyodonty, growing new teeth on the inside which migrated over time to replace the outer teeth. Unlike sharks however, who lose all of their old teeth, teeth in the rapidly growing dental battery would wear completely down and be reabsorbed by the renewing structure around it.[25]
The batteries were formed by the teeth growing fast and maturing early, to the point that the pulp cavity of individual teeth—usually filled with cells and connective tissue—were totally filled with dentine before it even erupted. The lack of pulp in the tooth post-eruption means that the tooth was essentially dead and able to be completely worn away through use, and replaced without the risk of exposing the normally sensitive dental pulp to infection and pain. While other dinosaurs, such as some ceratopsians and sauropods, also possessed dental batteries, they all evolved independently and differ in some form or function from those of hadrosaurs. This shows that some dinosaurs had evolved extremely sophisticated chewing capabilities.[24][25]
References
- Hendrickx, Christophe; Mateus, Octávio; Araújo, Ricardo; Choiniere, Jonah (2019). "The distribution of dental features in non-avian theropod dinosaurs: Taxonomic potential, degree of homoplasy, and major evolutionary trends". Palaeontologia Electronica. 22 (3): 1–110. doi:10.26879/820.
- Chinsamy, Anusuya; Rich, T.; Vickers-Rich, P. (1998-06-15). "Polar dinosaur bone histology". Journal of Vertebrate Paleontology. 18 (2): 385–390. doi:10.1080/02724634.1998.10011066. ISSN 0272-4634.
- Martin Sander, P.; Mateus, Octávio; Laven, Thomas; Knötschke, Nils (2006). "Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur". Nature. 441 (7094): 739–741. Bibcode:2006Natur.441..739M. doi:10.1038/nature04633. PMID 16760975. S2CID 4361820.
- Hübner, Tom R. (2012-01-06). "Bone Histology in Dysalotosaurus lettowvorbecki (Ornithischia: Iguanodontia) – Variation, Growth, and Implications". PLOS ONE. 7 (1): e29958. Bibcode:2012PLoSO...729958H. doi:10.1371/journal.pone.0029958. ISSN 1932-6203. PMC 3253128. PMID 22238683.
- Zhao, Qi; Benton, Michael J.; Sullivan, Corwin; Sander, P. Martin; Xu, Xing (2013-06-28). "Histology and postural change during the growth of the ceratopsian dinosaur Psittacosaurus lujiatunensis". Nature Communications. 4: ncomms3079. Bibcode:2013NatCo...4.2079Z. doi:10.1038/ncomms3079. PMID 23811819.
- Fong, R.K.M.; LeBlanc, A.R.; Berman, D.S.; Reisz, R.R. (2016). "Dental histology of Coelophysis bauri and the evolution of tooth attachment tissues in early dinosaurs". Journal of Morphology. 277 (7): 914–924. doi:10.1002/jmor.20545. PMID 27087142. S2CID 25916863.
- Brink, K. S.; Reisz, R. R.; LeBlanc, A. R. H.; Chang, R. S.; Lee, Y. C.; Chiang, C. C.; Huang, T.; Evans, D. C. (2015-07-28). "Developmental and evolutionary novelty in the serrated teeth of theropod dinosaurs". Scientific Reports. 5 (1): 12338. Bibcode:2015NatSR...512338B. doi:10.1038/srep12338. ISSN 2045-2322. PMC 4648475. PMID 26216577.
- Erickson, Gregory M.; Sidebottom, Mark A.; Kay, David I.; Turner, Kevin T.; Ip, Nathan; Norell, Mark A.; Sawyer, W. Gregory; Krick, Brandon A. (2015-06-01). "Wear biomechanics in the slicing dentition of the giant horned dinosaur Triceratops". Science Advances. 1 (5): e1500055. Bibcode:2015SciA....1E0055E. doi:10.1126/sciadv.1500055. ISSN 2375-2548. PMC 4640618. PMID 26601198.
- Erickson, Gregory M.; Krick, Brandon A.; Hamilton, Matthew; Bourne, Gerald R.; Norell, Mark A.; Lilleodden, Erica; Sawyer, W. Gregory (2012-10-05). "Complex Dental Structure and Wear Biomechanics in Hadrosaurid Dinosaurs". Science. 338 (6103): 98–101. Bibcode:2012Sci...338...98E. doi:10.1126/science.1224495. ISSN 0036-8075. PMID 23042891. S2CID 35186603.
- Sereno, Paul C.; Wilson, Jeffrey A.; Witmer, Lawrence M.; Whitlock, John A.; Maga, Abdoulaye; Ide, Oumarou; Rowe, Timothy A. (2007-11-21). "Structural Extremes in a Cretaceous Dinosaur". PLOS ONE. 2 (11): e1230. Bibcode:2007PLoSO...2.1230S. doi:10.1371/journal.pone.0001230. ISSN 1932-6203. PMC 2077925. PMID 18030355.
- LeBlanc, Aaron R. H.; Reisz, Robert R. (2013-09-04). "Periodontal Ligament, Cementum, and Alveolar Bone in the Oldest Herbivorous Tetrapods, and Their Evolutionary Significance". PLOS ONE. 8 (9): e74697. Bibcode:2013PLoSO...874697L. doi:10.1371/journal.pone.0074697. ISSN 1932-6203. PMC 3762739. PMID 24023957.
- LeBlanc, Aaron R. H.; Brink, Kirstin S.; Cullen, Thomas M.; Reisz, Robert R. (2017-09-29). "Evolutionary implications of tooth attachment versus tooth implantation: A case study using dinosaur, crocodilian, and mammal teeth". Journal of Vertebrate Paleontology. 37 (5): e1354006. doi:10.1080/02724634.2017.1354006. ISSN 0272-4634. S2CID 91066137.
- Hwang, S.H. (2005). "Phylogenetic Patterns of Enamel Microstructure in Dinosaur Teeth". Journal of Morphology. 266 (2): 208–240. doi:10.1002/jmor.10372. PMID 16163689. S2CID 9973691.
- Dauphin, Y. (1988). "L'email dentaire des Reptiles actuels et fossiles : repartition de la structure prismatique, son rôle, ses implications". Palaeontographica. A203: 171–184.
- Dauphin, Y.; Jaeger, J.J.; Osmolska, H (1988). "Enamel microstructure of ceratopsian teeth (Reptilia, Archosauria)". Geobios. 21 (3): 319–327. doi:10.1016/s0016-6995(88)80056-1. ISSN 0016-6995.
- Dauphin, Y. (1991). "Chemical composition of reptilian teeth - 2. implications for paleodiets". Palaeontographica. A219: 97–105.
- Bocherens, H.; Brinkman, D.B.; Dauphin, Y.; Mariotti, A. (1994). "Microstructural and geochemical investigations on Late Cretaceous archosaur teeth from Alberta, Canada". Canadian Journal of Earth Sciences. 31 (5): 783–792. Bibcode:1994CaJES..31..783B. doi:10.1139/e94-071. ISSN 0008-4077.
- Dauphin, Y.; Williams, C. T. (2007). "The chemical compositions of dentine and enamel from recent reptile and mammal teeth—variability in the diagenetic changes of fossil teeth". CrystEngComm. 9 (12): 1252–1261. doi:10.1039/b708985f. ISSN 1466-8033.
- Dauphin, Y.; Williams, C. T. (2008). "Chemical composition of enamel and dentine in modern reptile teeth". Mineralogical Magazine. 72 (1): 247–250. Bibcode:2008MinM...72..247D. doi:10.1180/minmag.2008.072.1.247. ISSN 0026-461X. S2CID 129102736.
- Erickson, G. M. (1996-12-10). "Incremental lines of von Ebner in dinosaurs and the assessment of tooth replacement rates using growth line counts". Proceedings of the National Academy of Sciences. 93 (25): 14623–14627. Bibcode:1996PNAS...9314623E. doi:10.1073/pnas.93.25.14623. ISSN 0027-8424. PMC 26184. PMID 8962103.
- Richman, Joy M.; Handrigan, Gregory R. (2011-04-01). "Reptilian tooth development". Genesis. 49 (4): 247–260. doi:10.1002/dvg.20721. ISSN 1526-968X. PMID 21309070.
- Wood, Sarah A.; Strait, David S.; Dumont, Elizabeth R.; Ross, Callum F.; Grosse, Ian R. (2011). "The effects of modeling simplifications on craniofacial finite element models: The alveoli (tooth sockets) and periodontal ligaments". Journal of Biomechanics. 44 (10): 1831–1838. doi:10.1016/j.jbiomech.2011.03.022. PMID 21592483.
- Porro, Laura B.; Holliday, Casey M.; Anapol, Fred; Ontiveros, Lupita C.; Ontiveros, Lolita T.; Ross, Callum F. (2011-08-01). "Free body analysis, beam mechanics, and finite element modeling of the mandible of Alligator mississippiensis". Journal of Morphology. 272 (8): 910–937. doi:10.1002/jmor.10957. ISSN 1097-4687. PMID 21567445. S2CID 10762324.
- LeBlanc, Aaron R. H.; Reisz, Robert R.; Evans, David C.; Bailleul, Alida M. (2016-07-28). "Ontogeny reveals function and evolution of the hadrosaurid dinosaur dental battery". BMC Evolutionary Biology. 16: 152. doi:10.1186/s12862-016-0721-1. ISSN 1471-2148. PMC 4964017. PMID 27465802.
- Bramble, Katherine; LeBlanc, Aaron R. H.; Lamoureux, Denis O.; Wosik, Mateusz; Currie, Philip J. (2017). "Histological evidence for a dynamic dental battery in hadrosaurid dinosaurs". Scientific Reports. 7 (1): 15787. Bibcode:2017NatSR...715787B. doi:10.1038/s41598-017-16056-3. PMC 5693932. PMID 29150664.