Eocyte hypothesis
The eocyte hypothesis in evolutionary biology proposes that the eukaryotes originated from a group of prokaryotes called eocytes (later classified as Thermoproteota, a group of archaea).[3] After his team at the University of California, Los Angeles discovered eocytes in 1984,[2] James A. Lake formulated the hypothesis as "eocyte tree" that proposed eukaryotes as part of archaea. Lake hypothesised the tree of life as having only two primary branches: Parkaryoates that include Bacteria and Archaea, and karyotes that comprise Eukaryotes and eocytes. Parts of this early hypothesis were revived in a newer two-domain system of biological classification which named the primary domains as Archaea and Bacteria.[4]
| Karyota | |
|---|---|
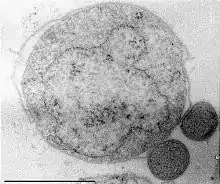 | |
| Ignicoccus hospitalis (and its symbiote Nanoarchaeum equitans) | |
| Scientific classification | |
| Domain: | |
| (unranked): | Karyota Lake (1988)[1] |
| Domain & Regnum | |
| |
| Synonyms | |
| |
Lake's hypothesis was based on an analysis of the structural components of ribosomes. It was largely ignored, being overshadowed by the three-domain system which relied on more precise genetic analysis. In 1990, Carl Woese and his colleagues proposed that cellular life consists of three domains – Eucarya, Bacteria, and Archaea – based on the ribosomal RNA sequences. The three-domain concept was widely accepted in genetics, and became the presumptive classification system for high-level taxonomy, and was promulgated in many textbooks.[5][6]
Resurgence of archaea research after the 2000s, using advanced genetic techniques, and later discoveries of new groups of archaea revived the eocyte hypothesis; consequently, the two-domain system has found wider acceptance.[7][8]
Description
In 1984, James A. Lake, Michael W. Clark, Eric Henderson, and Melanie Oakes of the University of California, Los Angeles described a new group of prokaryotic organisms designated as "a group of sulfur-dependent bacteria." Based on the structure and composition of their ribosomal subunits, they found that these organisms were different from other prokaryotes, bacteria and archaea, known at the time. They named them eocytes (for "dawn cells") and proposed a new biological kingdom Eocyta. According to this discovery, the tree of life is represented by four kingdoms, Archaebacteria, Eubacteria, Eukaryote and Eocyta.[2]
Following analyses of the rRNA sequences of the four groups, Lake concluded in 1988 that eukaryotes were closely related to eocytes such that the two groups constitute the same (monophyletic) group, meaning that eukaryotes originated from eocytes and not archaebacteria, as was generally assumed.[9] This was the establishment of the eocyte hypothesis.[3] In 1988, Lake proposed a systematic classification of all life forms into two taxonomic groups,[1] which he later mentioned as superkingdoms:[10]
- Karyotes (that include eukaryotes and proto-eukaryotic organisms such as eocytes)
- Parkaryotes (that consists of eubacteria and two groups of archaea known at the time, halobacteria and methanogens)[11]
Development and competition
Lake's classification was not widely recognised, but the eocyte hypothesis gained considerable attention after its introduction due to the interest in determining the origin of the eukaryotic cell.[12][13] However, the concept faced a problem because it was not known that eocytes, the main organism group on which the hypothesis was based, were archaea. For example, studies in the late 1980s and early 1990s still treated eocytes as separate group from archaea.[12][14][15] As Lake also argued, the rival hypothesis was called archaebacterial tree (as introduced by Carl Woese of the University of Illinois in 1987[16]) or archaebacterial theory, which (supposedly) stated that eukaryotes originated from archaea, and not eocytes.[10][17]
Due to such confusion, some studies appeared to invalidate the hypothesis. For example, Japanese scientists reported in 1990 their study on the elongation factors Tu(EF-Tu) and G(EF-G) from various organisms that showed that eukaryotes are most closely related to archaea (methanogen and halobacteria), and not eocytes.[15] Other studies also supported the eukaroyte-archaea relationship and rejected the eocyte hypotheses.[13][18][19] Ribosomal RNA sequencing in 1989 also opposed the eocyte tree as the origin of eukaryotes.[12]
Three-domain system
The most important blow to eocyte hypothesis and Lake's classification was the development of ribosomal RNA sequencing that became a reliable determinant in biological classification.[20][21] Introduced in 1977 by Carl Woese and George E. Fox[22] in classification, the technique indicated that archaea (with only methanogens known at the time) and bacteria were distinct groups of organisms. Two kingdoms, Archaebacteria (archaea) and Eubacteria (for bacteria) were established.[22] Based on further studies, Woese, Otto Kandler and Mark Wheelis introduced the concept of "domain" in 1990 as the highest level of biological classification, and proposed the three-domain system consisting of Eucarya, Bacteria and Archaea.[23] With it they classified eocytes as archaea under the phylum Crenarchaeota[24] (which was reclassified as Thermoproteota in 2021[25]).
The classification gradually gained acceptance and was recognised as "arguably the best-developed and most widely-accepted scientific hypotheses [with the five-kingdom classification] regarding the evolutionary history of life."[26] It became a scientific concept and general taxonomy in textbooks.[5][6] Although Lake continued to advocate his eocyte taxonomy and hypothesis instead of conceding that eocytes were archaea,[10][27] the hypothesis was largely neglected[28] and support of it waned in favour of the three-domain system.[3]
Archaeal studies
In addition to a Thermoproteota origin of eukaryotes, some studies have suggested that eukaryotes may also have originated in the Nitrososphaerota (formerly Thaumarchaeota).[3][29][30][31][32] A superphylum — TACK — has been proposed that includes the Nitrososphaerota, Thermoproteota, and other groups of archaea,[31] so that this superphylum may be related to the origin of eukaryotes. It is seen that eukaryotes share a large number of proteins with members of the TACK superphylum and that these complex archaea may have had rudimentary phagocytosis abilities to engulf bacteria.[24]
As a result of metagenomic analysis of material found nearby hydrothermal vents, another superphylum — Asgard — has been named and proposed to be more closely related to the original eukaryote and a sister group to TACK more recently. Asgard consists of phyla Lokiarchaeota (found first), Heimdallarchaeota (possibly related closest to eukaryotes) and others.[33][34]
Root of the eocyte tree

The eocyte tree root may be located in the RNA world; that is, the root organism may have been a ribocyte (also known as a ribocell). For cellular DNA and DNA handling, an "out of virus" scenario has been proposed: storing genetic information in DNA may have been an innovation performed by viruses and later handed over to ribocytes twice, once transforming them into bacteria and once transforming them into archaea.[35][36]
Although archaeal viruses are not as well-studied as bacterial phages, it is thought that dsDNA viruses led to the incorporation of the viral genome into archaeal genomes.[37] The transduction of genetic material through a viral vector led to an increase in complexity in the pre-eukaryotic cells.[38] All these findings do not change the eocyte tree as given here in principle, but examine a higher resolution of it.
Arguments against
Due to the similarities found between eukaryotes and both archaea and bacteria, it is thought that a major source of the genetic variation is through horizontal gene transfer.[39] Horizontal gene transfer explains why archaeal sequences are found in bacteria and bacterial sequences are found in archaea.[39] This could explain why elongation factors found in archaea and eukaryotes are so similar, the data currently out is obscured as horizontal gene transfer, vertical gene transfer, or endosymbiosis and could be behind the gene sequence similarity.[40] The eocyte hypothesis also has troubles due to the endosymbiotic theory, with the archaea being able to phagocytize bacteria for the formation of membrane-bound organelles.[41] It is thought that these ancestral prokaryotes began to have ectosymbiotic relationships with other prokaryotes and gradually engulfed these symbiotes through cell membrane protrusions.[42]
Although more recent data provides evidence in favour of the relationship between eukaryotes and Thermoproteota through the analysis of elongation factors, earlier experimentation with elongation factors provided evidence against such a relationship.[15] Hasegawa et al. uses these elongation factors to show that eukaryotes and archaebacteria are more closely related than archaebacteria and eubacteria than is explained in this two-tree system.[15]
Competing hypothesis
A competing hypothesis is that prokaryotes evolved towards thriving in higher temperatures to evade viruses through the thermoreductive hypothesis, however this does not account for the arising of eukaryotes and only takes into consideration the prokaryotic origins.[43] However decrease in complexity from a more complex origin is the basis of reductive evolution where a commensal relationship occurs, while this reduction explained in the thermoreduction hypothesis uses a parasitic relationship with viruses to explain the movement of complex pre-eukaryotes to a more harsh environment; that being ocean floor hydrothermal vents.[44]
Revival
Molecular studies
With advancements in genomics, the eocyte hypothesis experienced a revival beginning in the mid-2000s. As more archaeal genomes were sequenced, numerous genes coding for eukaryotic traits have been discovered in various archaean phyla, seemingly providing support for the eocyte hypothesis. Proteomics based research has also found supporting data with the use of elongation factor 1-α (eEF-1), a common housekeeping protein, to compare structural homology between eukaryotic and archaean lineages.[45] Furthermore, other proteins have been sequenced through proteomics with homologous structures in heat shock proteins found in both eukaryotes and archaea. The structure of these heat shock proteins were identified through X-ray crystallography to find the three dimensional structure of the proteins.[10] These proteins however have differing purposes as the eukaryote heat shock protein is a part of the T-complex while the archaeal heat shock protein is a molecular chaperone.[10] This creates an issue with the sequence homology that has been seen between 70 kilodalton heat shock proteins in eukaryotes and Gram-negative bacteria.[40]
Ribosome protein sequencing and phylogenetic analyses in 2004 showed that eukaryotes emerged from archaea.[46][47] Phylogenomic analysis in 2007 also pointed to the origin of eukaryotes specifically from the Thermoplasmatales.[48] The so-called "eukaryotic signature proteins" actin (cytoskeletal microfilament involved in cell motility), tubulin (component of the large cytoskeleton, microtubule) and the ubiquitin system (protein degradation and recycling)[7][49] which are thought to be unique to eukaryotes were found in TACK (comprising the phyla Thaumarchaeota, Aigarchaeota, Crenarchaeota and Korarchaeota) archaea but not in other archaea. These indicate that eukaryotes can be merged into archaea.[24]
Discovery of Asgards
Asgard, described as "eukaryote-like archaea",[50] were discovered in 2012.[51][52] The first known Asgards called Lokiarchaeota contain more eukaryotic protein-genes than the TACK group that supported the merging of eukaryote–archaea grouping, meaning a single domain of Archaea.[53][54] Phylogenomic studies indicated that Heimdallarchaeota, another group of Asgards, are the closest relatives of eukaryotes.[34][55][56] A new group of Asgard described in 2021 (named Wukongarchaeota) are also among the eukaryotic roots.[57] Another new Asgard reported in 2022, named Njordarchaeota, is related to the Heimdallarchaeota–Wukongarchaeota branch and is possibly the origin group for eukaryotes.[58]
The Asgards contain at least 80 genes for eukaryotic signature proteins.[59] In addition to actin, tubulin, ubiquitin and ESCRT proteins found in TACK archaea, Asgards contain functional genes for several other eukaryotic proteins such as profilins,[60] ubiquitin system (E1-like, E2-like and small-RING finger (srfp) proteins),[61] membrane-trafficking systems (such as Sec23/24 and TRAPP domains), variety of small GTPases[62] (including Gtr/Rag family GTPase orthologues[63]), and gelsolins.[64]
The two-domain system
As more archaea were later discovered and better genetic analysis were available, it was realised that the three-domain concept might not have represented the correct origin of eukaryotes.[65][32] Ford Doolittle, then at the Dalhousie University, Canada, wrote in 2020:
- "[The] three-domain tree wrongly represents evolutionary relationships, presenting a misleading view about how eukaryotes evolved from prokaryotes. The three-domain tree does recognize a specific archaeal–eukaryotic affinity, but it would have the latter arising independently, not from within, the former."[66]
This is because research since the early 2000s have revealed two important issues: eukaryotes originated within Archaea, and a new group of archaea called Asgards represent the root of eukaryotes.[67][68] This led to the rebirth of the eocyte hypothesis and development of the two-domain system.[3]
Discoveries of eukaryotic signature proteins in TACK and Asgard archaea support the notion that eukaryotes evolved from archaea. Discoveries of more Asgards and better understanding of their nature indicate that they are the likely root of eukaryotes and are considered the strong "evidence of the Eocyte hypothesis."[67] Although these facts do not completely rule out the three-domain concept,[50] they generally strengthened the two-domain system.[7][56][57]
References
- Lake JA (January 1988). "Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences". Nature. 331 (6152): 184–186. Bibcode:1988Natur.331..184L. doi:10.1038/331184a0. PMID 3340165. S2CID 4368082.
- Lake JA, Henderson E, Oakes M, Clark MW (June 1984). "Eocytes: a new ribosome structure indicates a kingdom with a close relationship to eukaryotes". Proceedings of the National Academy of Sciences of the United States of America. 81 (12): 3786–3790. Bibcode:1984PNAS...81.3786L. doi:10.1073/pnas.81.12.3786. PMC 345305. PMID 6587394.
- Archibald JM (December 2008). "The eocyte hypothesis and the origin of eukaryotic cells". Proceedings of the National Academy of Sciences of the United States of America. 105 (51): 20049–20050. Bibcode:2008PNAS..10520049A. doi:10.1073/pnas.0811118106. PMC 2629348. PMID 19091952.
- Zhou Z, Liu Y, Li M, Gu JD (April 2018). "Two or three domains: a new view of tree of life in the genomics era". Applied Microbiology and Biotechnology. 102 (7): 3049–3058. doi:10.1007/s00253-018-8831-x. PMID 29484479. S2CID 3541409.
- Peirce SK (February 1999). "Three domains, not five kingdoms: A phylogenetic classification system". The American Biology Teacher. 61 (2): 132–136. doi:10.2307/4450634. ISSN 0002-7685. JSTOR 4450634.
- Forterre, P. (2015). "The universal tree of life: An update". Frontiers in Microbiology. 6: 717. doi:10.3389/fmicb.2015.00717. PMC 4508532. PMID 26257711.
- Nobs SJ, MacLeod FI, Wong HL, Burns BP (May 2022). "Eukarya the chimera: Eukaryotes, a secondary innovation of the two domains of life?". Trends in Microbiology. 30 (5): 421–431. doi:10.1016/j.tim.2021.11.003. PMID 34863611. S2CID 244823103.
- Williams TA, Cox CJ, Foster PG, Szöllősi GJ, Embley TM (January 2020). "Phylogenomics provides robust support for a two-domains tree of life". Nature Ecology & Evolution. 4 (1): 138–147. doi:10.1038/s41559-019-1040-x. PMC 6942926. PMID 31819234.
- Lake JA (1987). "Prokaryotes and archaebacteria are not monophyletic: rate invariant analysis of rRNA genes indicates that eukaryotes and eocytes form a monophyletic taxon". Cold Spring Harbor Symposia on Quantitative Biology. 52: 839–846. doi:10.1101/SQB.1987.052.01.091. PMID 3454292.
- Rivera MC, Lake JA (July 1992). "Evidence that eukaryotes and eocyte prokaryotes are immediate relatives". Science. 257 (5066): 74–76. Bibcode:1992Sci...257...74R. doi:10.1126/science.1621096. PMID 1621096.
- Lake JA (1991). "Tracing origins with molecular sequences: metazoan and eukaryotic beginnings". Trends in Biochemical Sciences. 16 (2): 46–50. doi:10.1016/0968-0004(91)90020-V. PMID 1858129.
- Gouy M, Li WH (May 1989). "Phylogenetic analysis based on rRNA sequences supports the archaebacterial rather than the eocyte tree". Nature. 339 (6220): 145–147. Bibcode:1989Natur.339..145G. doi:10.1038/339145a0. PMID 2497353. S2CID 4315004.
- Klenk HP, Palm P, Zillig W (1991). "A monophyletic holophyletic archaeal domain versus the 'eocyte tree'". Trends in Biochemical Sciences. 16 (8): 288–289. doi:10.1016/0968-0004(91)90118-F. PMID 1796995.
- Gogarten JP, Kibak H, Dittrich P, Taiz L, Bowman EJ, Bowman BJ, et al. (September 1989). "Evolution of the vacuolar H+-ATPase: implications for the origin of eukaryotes". Proceedings of the National Academy of Sciences of the United States of America. 86 (17): 6661–6665. Bibcode:1989PNAS...86.6661G. doi:10.1073/pnas.86.17.6661. PMC 297905. PMID 2528146.
- Hasegawa M, IwABE N, Mukohata Y, Miyata T (1990). "Close evolutionary relatedness of archaebacteria, Methanococcus and Halobacterium, to eukaryotes demonstrated by composite phylogenetic trees of elongation factors EF-Tu and EF-G: Eocyte tree is unlikely". The Japanese Journal of Genetics. 65 (3): 109–114. doi:10.1266/jjg.65.109. ISSN 0021-504X.
- Woese CR (June 1987). "Bacterial evolution". Microbiological Reviews. 51 (2): 221–271. doi:10.1128/mr.51.2.221-271.1987. PMC 373105. PMID 2439888.
- Achenbach-Richter L, Gupta R, Zillig W, Woese CR (1988). "Rooting the archaebacterial tree: the pivotal role of Thermococcus celer in archaebacterial evolution". Systematic and Applied Microbiology. 10 (3): 231–240. doi:10.1016/S0723-2020(88)80007-9. PMID 11542150.
- Olsen GJ, Woese CR (January 1989). "A brief note concerning archaebacterial phylogeny". Canadian Journal of Microbiology. 35 (1): 119–123. doi:10.1139/m89-018. PMID 2497936.
- Puglisi JD, Tan R, Calnan BJ, Frankel AD, Williamson JR (July 1992). "Conformation of the TAR RNA-arginine complex by NMR spectroscopy". Science. 257 (5066): 76–80. Bibcode:1992Sci...257...76P. doi:10.1126/science.1621097. PMID 1621097.
- Ludwig W, Schleifer KH (October 1994). "Bacterial phylogeny based on 16S and 23S rRNA sequence analysis". FEMS Microbiology Reviews. 15 (2–3): 155–173. doi:10.1111/j.1574-6976.1994.tb00132.x. PMID 7524576. S2CID 8950474.
- Ludwig W, Strunk O, Klugbauer S, Klugbauer N, Weizenegger M, Neumaier J, et al. (April 1998). "Bacterial phylogeny based on comparative sequence analysis". Electrophoresis. 19 (4): 554–568. doi:10.1002/elps.1150190416. PMID 9588802. S2CID 28196433.
- Woese CR, Fox GE (November 1977). "Phylogenetic structure of the prokaryotic domain: The primary kingdoms". Proceedings of the National Academy of Sciences of the United States of America. 74 (11): 5088–5090. Bibcode:1977PNAS...74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104. PMID 270744.
- Woese CR, Kandler O, Wheelis ML (June 1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". Proceedings of the National Academy of Sciences of the United States of America. 87 (12): 4576–4579. Bibcode:1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC 54159. PMID 2112744.
- Koonin EV (September 2015). "Origin of eukaryotes from within archaea, archaeal eukaryome and bursts of gene gain: eukaryogenesis just made easier?". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1678): 20140333. doi:10.1098/rstb.2014.0333. PMC 4571572. PMID 26323764.
- Oren A, Garrity GM (October 2021). "Valid publication of the names of forty-two phyla of prokaryotes". International Journal of Systematic and Evolutionary Microbiology. 71 (10): Online. doi:10.1099/ijsem.0.005056. PMID 34694987. S2CID 239887308.
- Case E (2008). "Teaching Taxonomy: How Many Kingdoms?". The American Biology Teacher. 70 (8): 472–477. doi:10.1662/0002-7685(2008)70[472:TTHMK]2.0.CO;2. S2CID 198969439.
- Lake JA (February 1994). "Reconstructing evolutionary trees from DNA and protein sequences: paralinear distances". Proceedings of the National Academy of Sciences of the United States of America. 91 (4): 1455–1459. Bibcode:1994PNAS...91.1455L. doi:10.1073/pnas.91.4.1455. PMC 43178. PMID 8108430.
- Foster PG, Cox CJ, Embley TM (August 2009). "The primary divisions of life: a phylogenomic approach employing composition-heterogeneous methods". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 364 (1527): 2197–2207. doi:10.1098/rstb.2009.0034. PMC 2873002. PMID 19571240.
- Kelly S, Wickstead B, Gull K (April 2011). "Archaeal phylogenomics provides evidence in support of a methanogenic origin of the Archaea and a thaumarchaeal origin for the eukaryotes" (PDF). Proceedings. Biological Sciences. 278 (1708): 1009–1018. doi:10.1098/rspb.2010.1427. PMC 3049024. PMID 20880885. Archived from the original (PDF) on 3 March 2016. Retrieved 5 October 2012.
- Poole AM, Neumann N (January 2011). "Reconciling an archaeal origin of eukaryotes with engulfment: a biologically plausible update of the Eocyte hypothesis". Research in Microbiology. 162 (1): 71–76. doi:10.1016/j.resmic.2010.10.002. PMID 21034814.
- Guy L, Ettema TJ (December 2011). "The archaeal 'TACK' superphylum and the origin of eukaryotes". Trends in Microbiology. 19 (12): 580–587. doi:10.1016/j.tim.2011.09.002. PMID 22018741.
- Cox CJ, Foster PG, Hirt RP, Harris SR, Embley TM (December 2008). "The archaebacterial origin of eukaryotes". Proceedings of the National Academy of Sciences of the United States of America. 105 (51): 20356–20361. Bibcode:2008PNAS..10520356C. doi:10.1073/pnas.0810647105. PMC 2629343. PMID 19073919.
- Eme L, Spang A, Lombard J, Stairs CW, Ettema TJ (November 2017). "Archaea and the origin of eukaryotes". Nature Reviews. Microbiology. 15 (12): 711–723. doi:10.1038/nrmicro.2017.133. PMID 29123225. S2CID 8666687.
- Zaremba-Niedzwiedzka K, Caceres EF, Saw JH, Bäckström D, Juzokaite L, Vancaester E, et al. (January 2017). "Asgard archaea illuminate the origin of eukaryotic cellular complexity" (PDF). Nature. 541 (7637): 353–358. Bibcode:2017Natur.541..353Z. doi:10.1038/nature21031. OSTI 1580084. PMID 28077874. S2CID 4458094.
- Forterre P, Krupovic M (2012). "The Origin of Virions and Virocells: The Escape Hypothesis Revisited". In Witzany G (ed.). Viruses: Essential Agents of Life. pp. 43–60. doi:10.1007/978-94-007-4899-6_3. ISBN 978-94-007-4898-9.
- Forterre, P. (August 2017). "Die wahre Natur der Viren" [The real nature of viruses]. Evolution. Spektrum (in German). p. 37. Retrieved 19 July 2017.
- Zillig W, Prangishvilli D, Schleper C, Elferink M, Holz I, Albers S, et al. (May 1996). "Viruses, plasmids and other genetic elements of thermophilic and hyperthermophilic Archaea". FEMS Microbiology Reviews. 18 (2–3): 225–236. doi:10.1016/0168-6445(96)00014-9. PMID 8639330.
- Forterre, P. (2013). "The common ancestor of archaea and eukarya was not an archaeon". Archaea. 2013: 372396. doi:10.1155/2013/372396. PMC 3855935. PMID 24348094.
- Leonard CJ, Aravind L, Koonin EV (October 1998). "Novel families of putative protein kinases in bacteria and archaea: evolution of the "eukaryotic" protein kinase superfamily". Genome Research. 8 (10): 1038–1047. doi:10.1101/gr.8.10.1038. PMID 9799791.
- Gupta RS, Singh B (December 1994). "Phylogenetic analysis of 70 kD heat shock protein sequences suggests a chimeric origin for the eukaryotic cell nucleus". Current Biology. 4 (12): 1104–1114. doi:10.1016/s0960-9822(00)00249-9. PMID 7704574. S2CID 2568264.
- Davidov Y, Jurkevitch E (July 2009). "Predation between prokaryotes and the origin of eukaryotes". BioEssays. 31 (7): 748–757. doi:10.1002/bies.200900018. PMID 19492355. S2CID 8796311.
- Baum DA, Baum B (October 2014). "An inside-out origin for the eukaryotic cell". BMC Biology. 12: 76. doi:10.1186/s12915-014-0076-2. PMC 4210606. PMID 25350791.
- Forterre, P. (April 1995). "Thermoreduction, a hypothesis for the origin of prokaryotes". Comptes Rendus de l'Académie des Sciences. III, Sciences de la Vie. 318 (4): 415–422. PMID 7648354.
- Martin WF, Garg S, Zimorski V (September 2015). "Endosymbiotic theories for eukaryote origin". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1678): 20140330. doi:10.1098/rstb.2014.0330. PMC 4571569. PMID 26323761.
- Roger AJ, Sandblom O, Doolittle WF, Philippe H (February 1999). "An evaluation of elongation factor 1 alpha as a phylogenetic marker for eukaryotes". Molecular Biology and Evolution. 16 (2): 218–233. doi:10.1093/oxfordjournals.molbev.a026104. PMID 10028289.
- Vishwanath P, Favaretto P, Hartman H, Mohr SC, Smith TF (December 2004). "Ribosomal protein-sequence block structure suggests complex prokaryotic evolution with implications for the origin of eukaryotes". Molecular Phylogenetics and Evolution. 33 (3): 615–625. doi:10.1016/j.ympev.2004.07.003. PMID 15522791.
- Rivera MC, Lake JA (September 2004). "The ring of life provides evidence for a genome fusion origin of eukaryotes". Nature. 431 (7005): 152–155. Bibcode:2004Natur.431..152R. doi:10.1038/nature02848. PMID 15356622. S2CID 4349149.
- Pisani D, Cotton JA, McInerney JO (August 2007). "Supertrees disentangle the chimerical origin of eukaryotic genomes". Molecular Biology and Evolution. 24 (8): 1752–1760. doi:10.1093/molbev/msm095. PMID 17504772.
- Yutin N, Wolf MY, Wolf YI, Koonin EV (February 2009). "The origins of phagocytosis and eukaryogenesis". Biology Direct. 4: 9. doi:10.1186/1745-6150-4-9. PMC 2651865. PMID 19245710.
- Fournier GP, Poole AM (2018). "A Briefly Argued Case That Asgard Archaea Are Part of the Eukaryote Tree". Frontiers in Microbiology. 9: 1896. doi:10.3389/fmicb.2018.01896. PMC 6104171. PMID 30158917.
- Jorgensen SL, Hannisdal B, Lanzén A, Baumberger T, Flesland K, Fonseca R, et al. (October 2012). "Correlating microbial community profiles with geochemical data in highly stratified sediments from the Arctic Mid-Ocean Ridge". Proceedings of the National Academy of Sciences of the United States of America. 109 (42): E2846–E2855. doi:10.1073/pnas.1207574109. PMC 3479504. PMID 23027979.
- Jørgensen SL, Thorseth IH, Pedersen RB, Baumberger T, Schleper C (4 October 2013). "Quantitative and phylogenetic study of the Deep Sea Archaeal Group in sediments of the Arctic mid-ocean spreading ridge". Frontiers in Microbiology. 4: 299. doi:10.3389/fmicb.2013.00299. PMC 3790079. PMID 24109477.
- Da Cunha V, Gaia M, Gadelle D, Nasir A, Forterre P (June 2017). "Lokiarchaea are close relatives of Euryarchaeota, not bridging the gap between prokaryotes and eukaryotes". PLOS Genetics. 13 (6): e1006810. doi:10.1371/journal.pgen.1006810. PMC 5484517. PMID 28604769.
- da Cunha V, Gaïa M, Forterre P (2022). "The expanding Asgard archaea and their elusive relationships with Eukarya". mLife. 1 (1): 3–12. doi:10.1002/mlf2.12012. S2CID 247689333.
- Spang A, Eme L, Saw JH, Caceres EF, Zaremba-Niedzwiedzka K, Lombard J, et al. (March 2018). "Asgard archaea are the closest prokaryotic relatives of eukaryotes". PLOS Genetics. 14 (3): e1007080. doi:10.1371/journal.pgen.1007080. PMC 5875740. PMID 29596421.
- Williams TA, Cox CJ, Foster PG, Szöllősi GJ, Embley TM (January 2020). "Phylogenomics provides robust support for a two-domains tree of life". Nature Ecology & Evolution. 4 (1): 138–147. doi:10.1038/s41559-019-1040-x. PMC 6942926. PMID 31819234.
- Liu Y, Makarova KS, Huang WC, Wolf YI, Nikolskaya AN, Zhang X, et al. (May 2021). "Expanded diversity of Asgard archaea and their relationships with eukaryotes". Nature. 593 (7860): 553–557. Bibcode:2021Natur.593..553L. doi:10.1038/s41586-021-03494-3. PMID 33911286. S2CID 233447651.
- Xie R, Wang Y, Huang D, Hou J, Li L, Hu H, et al. (April 2022). "Expanding Asgard members in the domain of Archaea sheds new light on the origin of eukaryotes". Science China Life Sciences. 65 (4): 818–829. doi:10.1007/s11427-021-1969-6. PMID 34378142. S2CID 236977853.
- López-García P, Moreira D (April 2020). "Cultured Asgard Archaea Shed Light on Eukaryogenesis". Cell. 181 (2): 232–235. doi:10.1016/j.cell.2020.03.058. PMID 32302567. S2CID 215798394.
- Akıl C, Robinson RC (October 2018). "Genomes of Asgard archaea encode profilins that regulate actin". Nature. 562 (7727): 439–443. Bibcode:2018Natur.562..439A. doi:10.1038/s41586-018-0548-6. PMID 30283132. S2CID 52917038.
- Hennell James R, Caceres EF, Escasinas A, Alhasan H, Howard JA, Deery MJ, et al. (October 2017). "Functional reconstruction of a eukaryotic-like E1/E2/(RING) E3 ubiquitylation cascade from an uncultured archaeon". Nature Communications. 8 (1): 1120. Bibcode:2017NatCo...8.1120H. doi:10.1038/s41467-017-01162-7. PMC 5654768. PMID 29066714.
- Spang A, Saw JH, Jørgensen SL, Zaremba-Niedzwiedzka K, Martijn J, Lind AE, et al. (May 2015). "Complex archaea that bridge the gap between prokaryotes and eukaryotes". Nature. 521 (7551): 173–179. Bibcode:2015Natur.521..173S. doi:10.1038/nature14447. PMC 4444528. PMID 25945739.
- Klinger CM, Spang A, Dacks JB, Ettema TJ (June 2016). "Tracing the Archaeal Origins of Eukaryotic Membrane-Trafficking System Building Blocks". Molecular Biology and Evolution. 33 (6): 1528–1541. doi:10.1093/molbev/msw034. PMID 26893300.
- Akıl C, Tran LT, Orhant-Prioux M, Baskaran Y, Manser E, Blanchoin L, Robinson RC (August 2020). "Insights into the evolution of regulated actin dynamics via characterization of primitive gelsolin/cofilin proteins from Asgard archaea". Proceedings of the National Academy of Sciences of the United States of America. 117 (33): 19904–19913. Bibcode:2020PNAS..11719904A. doi:10.1073/pnas.2009167117. PMC 7444086. PMID 32747565.
- Zhou Z, Liu Y, Li M, Gu JD (April 2018). "Two or three domains: a new view of tree of life in the genomics era". Applied Microbiology and Biotechnology. 102 (7): 3049–3058. doi:10.1007/s00253-018-8831-x. PMID 29484479. S2CID 3541409.
- Doolittle WF (February 2020). "Evolution: Two Domains of Life or Three?". Current Biology. 30 (4): R177–R179. doi:10.1016/j.cub.2020.01.010. PMID 32097647.
- MacLeod F, Kindler GS, Wong HL, Chen R, Burns BP (2019). "Asgard archaea: Diversity, function, and evolutionary implications in a range of microbiomes". AIMS Microbiology. 5 (1): 48–61. doi:10.3934/microbiol.2019.1.48. PMC 6646929. PMID 31384702.
- Jüttner M, Ferreira-Cerca S (April 2022). "Looking through the Lens of the Ribosome Biogenesis Evolutionary History: Possible Implications for Archaeal Phylogeny and Eukaryogenesis". Molecular Biology and Evolution. 39 (4): msac054. doi:10.1093/molbev/msac054. PMC 8997704. PMID 35275997.