Ephaptic coupling
Ephaptic coupling is a form of communication within the nervous system and is distinct from direct communication systems like electrical synapses and chemical synapses. It may refer to the coupling of adjacent (touching) nerve fibers caused by the exchange of ions between the cells, or it may refer to coupling of nerve fibers as a result of local electric fields.[1] In either case ephaptic coupling can influence the synchronization and timing of action potential firing in neurons. Myelination is thought to inhibit ephaptic interactions.[2]
History and etymology
The idea that the electrical activity generated by nervous tissue may influence the activity of surrounding nervous tissue is one that dates back to the late 19th century. Early experiments, like those by Emil du Bois-Reymond,[3] demonstrated that the firing of a primary nerve may induce the firing of an adjacent secondary nerve (termed "secondary excitation"). This effect was not quantitatively explored, however, until experiments by Katz and Schmitt[4] in 1940, when the two explored the electric interaction of two adjacent limb nerves of the crab Carcinus maenas. Their work demonstrated that the progression of the action potential in the active axon caused excitability changes in the inactive axon. These changes were attributed to the local currents that form the action potential. For example, the currents that caused the depolarization (excitation) of the active nerve caused a corresponding hyperpolarization (depression) of the adjacent resting fiber. Similarly, the currents that caused repolarization of the active nerve caused slight depolarization in the resting fiber. Katz and Schmitt also observed that stimulation of both nerves could cause interference effects. Simultaneous action potential firing caused interference and resulted in decreased conduction velocity, while slightly offset stimulation resulted in synchronization of the two impulses.
In 1941 Angélique Arvanitaki[5] explored the same topic and proposed the usage of the term "ephapse" (from the Greek ephapsis and meaning "to touch") to describe this phenomenon and distinguish it from synaptic transmission. Over time the term ephaptic coupling has come to be used not only in cases of electric interaction between adjacent elements, but also more generally to describe the effects induced by any field changes along the cell membrane.[6]
Mechanism and effects
Role in excitation and inhibition
The early work performed by Katz and Schmitt demonstrated that ephaptic coupling between the two adjacent nerves was insufficient to stimulate an action potential in the resting nerve. Under ideal conditions the maximum depolarization observed was approximately 20% of the threshold stimulus.[4] However, conditions can be manipulated in such a way that the action potential from one neuron can be spread to a neighboring neuron. This was accomplished in one study in two experimental conditions: increased calcium concentrations, which lowered the threshold potential, or by submerging the axons in mineral oil, which increased resistance. While these manipulations do not reflect normal conditions, they do highlight the mechanisms behind ephaptic excitation.[7]
Ephaptic coupling has also been found to play an important role in inhibition of neighboring neurons. Depending on the location and identity of the neurons, various mechanisms have been found to underlie ephaptic inhibition. In one study, newly excited neighboring neurons interfered with already sustained currents, thus lowering the extracellular potential and depolarizing the neuron in relation to its surrounding environment, effectively inhibiting the action potential's propagation.[8]
Role in synchronization and timing
Studies of ephaptic coupling have also focused on its role in the synchronization and timing of action potentials in neurons. In the simpler case of adjacent fibers that experience simultaneous stimulation the impulse is slowed because both fibers are limited to exchange ions solely with the interstitial fluid (increasing the resistance of the nerve). Slightly offset impulses (conduction velocities differing by less than 10%) are able to exchange ions constructively and the action potentials propagate slightly out of phase at the same velocity.
More recent research, however, has focused on the more general case of electric fields that affect a variety of neurons. It has been observed that local field potentials in cortical neurons can serve to synchronize neuronal activity.[9] Although the mechanism is unknown, it is hypothesized that neurons are ephaptically coupled to the frequencies of the local field potential. This coupling may effectively synchronize neurons into periods of enhanced excitability (or depression) and allow for specific patterns of action potential timing (often referred to as spike timing). This effect has been demonstrated and modeled in a variety of cases.[10][11]
A hypothesis or explanation behind the mechanism is "one-way", "master-slave", or "unidirectional synchronization" effect as mathematical and fundamental property of non-linear dynamic systems (oscillators like neurons) to synchronize under certain criteria. Such phenomenon was proposed and predicted to be possible between two HR neurons, since 2010 in simulations and modeling work by Hrg.[12] It was also shown that such unidirectional synchronization or copy/paste transfer of neural dynamics from master to slave neurons, could be exhibited in different ways. Hence the phenomenon is of not only fundamental interest but also applied one from treating epilepsy to novel learning systems. A study in July 2023 found that mathematical models of ephaptic coupling predicted in vivo data of neural activity.[13] The authors likened the electric field to a conductor of an orchestra and neurons to the musicians. Then the field,like the conductor, listens to the music and guides the musicians accordingly. In an opinion paper, they also suggested that not only neurons but other parts of the cytoskeleton generate electromagnetic fields that influence individual neurons, and called this cytoelectric coupling.[14] Synchronization of neurons is in principle unwanted behavior, as brain would have zero information or be simply a bulb if all neurons would synchronize. Hence it is a hypothesis that neurobiology and evolution of brain coped with ways of preventing such synchronous behavior on large scale, using it rather in other special cases.
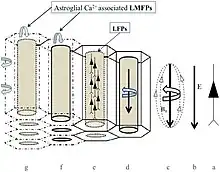
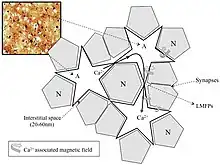
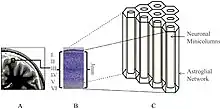
As models of brain function using only neuronal and gap junction connections fail to explain its complexity ephaptic coupling is being added more to the equation to try and explain the isopotentiality of cortical astrocytes to maintain the bioelectromagnetic crosstalk between neurons and astrocytes in the neocortex.[15]
Examples
Cardiac tissue
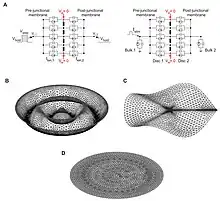
The electrical conduction system of the heart has been robustly established. However, newer research has been challenging some of the previously accepted models. The role of ephaptic coupling in cardiac cells is becoming more apparent. One author even goes so far as to say, “While previously viewed as a possible alternative to electrotonic coupling, ephaptic coupling has since come to be viewed as operating in tandem with gap junctions, helping sustain conduction when gap junctional coupling is compromised.”[16] Ephaptic interactions among cardiac cells help fill in the gaps that electrical synapses alone cannot account for. The proximity of sodium channels to gap junction plaques has been shown to relate to their effectiveness in driving ephaptic coupling action potential transmission.[17] There are also a number of mathematical models that now incorporate ephaptic coupling into predictions about electrical conductance in the heart.[18] Experimental work suggests that sodium channel-rich nanodomains located at sites of close contact between cardiac myocytes may constitute functional units of ephaptic coupling and selective disruption of these nanodomains resulted in arrhythmogenic conduction slowing, suggesting an important role for ephaptic coupling in the heart.[19] Potential ephaptic connections are now being considered in heart therapeutics.[20][21]
Epilepsy and seizures
Epileptic seizures occur when there is synchrony of electrical waves in the brain. Knowing the role that ephaptic coupling plays in maintaining synchrony in electrical signals, it makes sense to look for ephaptic mechanisms in this type of pathology. One study suggested that cortical cells represent an ideal place to observe ephaptic coupling due to the tight packing of axons, which allows for interactions between their electrical fields. They tested the effects of changing extracellular space (which affects local electrical fields) and found that one can block epileptic synchronization independent of chemical synapse manipulation simply by increasing the space between cells.[22] Later, a model was created to predict this phenomenon and showed scenarios with greater extracellular spacing that effectively blocked epileptic synchronization in the brain.[23]
Olfactory system in the brain
Neurons in the olfactory system are unmyelinated and densely packed and thus the often small effects of ephaptic coupling are more easily seen. A number of studies have shown how inhibition among neurons in the olfactory system work to fine-tune integration of signals in response to odor. This inhibition has been shown to occur from changes in electrical potentials alone.[8] The addition of ephaptic coupling to olfactory neuron models adds further support to the "dedicated-line" model in which each olfactory receptor sends its signal to one neuron. The inhibition due to ephaptic coupling would help account for the integration of signals that gives rise to more nuanced perception of smells.[24]
Synapses
Synapses are typically formed where two neurons come in very close proximity. Ephaptic coupling may therefore arise across the synaptic cleft. Prominent examples are the fish Mauthner cell,[25] the chick ciliary ganglion,[26] the mammalian cerebellar basket synapse,[27] and the mammalian, developing calyx of Held synapse.[28]
Mathematical models
Due to the very small electrical fields produced by neurons, mathematical models are often used in order to test a number of manipulations. Cable theory is one of the most important mathematical equations in neuroscience.[29] It calculates electric current using capacitance and resistance as variables and has been the main basis for many predictions about ephaptic coupling in neurons. However, many authors have worked to create more refined models in order to more accurately represent the environments of the nervous system. For example, many authors have proposed models for cardiac tissue that includes additional variables that account for the unique structure and geometry of cardiac cells [18] varying scales of size,[30] or three-dimensional electrodiffusion.[31]
Animal models
Squid giant axons
In 1978, basic tests were being conducted on squid giant axons in order to find evidence of ephaptic events. It was shown that an action potential of one axon could be propagated to a neighboring axon. The level of transmission varied, from subthreshold changes to initiation of an action potential in a neighboring cell, but in all cases, it was apparent that there are implications of ephaptic coupling that are of physiological importance.[7]
Rat spinal cord and medulla
One study tested the effects of ephaptic coupling by using both neurotransmitter antagonists to block chemical synapses and gap junction blockers to block electrical synapses. It was found that rhythmic electrical discharge associated with fetal neurons in the rat spinal cord and medulla was still sustained. This suggests that connections between the neurons still exist and work to spread signals even without traditional synapses. These findings support a model in which ephaptic coupling works alongside canonical synapses to propagate signals across neuronal networks.[32]
Rat Purkinje cells of the cerebellum
One of the few known cases of a functional system in which ephaptic coupling is responsible for an observable physiological event is in the Purkinje cells of the rat cerebellum.[27] It was demonstrated in this study that the basket cells which encapsulate some regions of Purkinje fibers can cause inhibitory effects on the Purkinje cells. The firing of these basket cells, which occurs more rapidly than in the Purkinje cells, draws current across the Purkinje cell and generates a passive hyperpolarizing potential which inhibits the activity of the Purkinje cell. Although the exact functional role of this inhibition is still unclear, it may well have a synchronizing effect in the Purkinje cells as the ephaptic effect will limit the firing time.
A similar ephaptic effect has been studied in the Mauthner cells of teleosts.[33]
Skepticism
While the idea of non-synaptic interactions between neurons has existed since the 19th century, there has historically been a lot of skepticism in the field of neuroscience. Many people believed that the micro electrical fields produced by the neurons themselves were so small that they were negligible.[24] While many supporters of the ephaptic coupling theory have been trying to prove its existence through experiments that block both chemical and electrical synapses, still some opponents in the field express caution. For example, in 2014, one scientist published a review that presents his skepticism on the idea of ephaptic coupling, saying “The agreement between their simulations and Poelzing’s data is impressive, but I will need a more definitive experimental confirmation before I can embrace the ephaptic hypothesis.” [34] He bases his caution in wanting more distinction between gap junctions' propagation of charge and true ephaptic coupling. Whether it is a true lack of evidence or simply obstinance in the face of change, many in the field are still not entirely convinced there is unambiguous evidence of ephaptic coupling.
Research continues and in 2018 what appears to be a definitive experiment was published.[35] A process of slow wave propagation in the brain of about 0.1m per second has been shown in vivo. Slow wave propagation was also recorded by inserting two electrodes into two ends of a piece of mouse brain taken from a freshly killed mouse. The tissue was then cut between the two electrode positions, severing any possible link by neurons or gap junctions between the two electrodes. The slow wave continued to propagate and could only be stopped by separating the two pieces of tissue with a 400-micron gap. Assuming the reproducibility of this experiment skeptics of ephaptic coupling should be satisfied it is a real phenomenon and investigations will focus now on its mechanisms and role rather than its existence.
See also
- Saltatory conduction
- Electroencephalography
- Spike-field coherence
- NeuroElectroDynamics
- Cable theory
- Local field potential
References
- Aur D., Jog, MS. (2010) Neuroelectrodynamics: Understanding the brain language, IOS Press, doi:10.3233/978-1-60750-473-3-i
- Hartline DK (May 2008). "What is myelin?". Neuron Glia Biology. 4 (2): 153–63. doi:10.1017/S1740925X09990263. PMID 19737435.
- Wilhelm Biedermann (1898). Electro-physiology. Macmillan. pp. 270–.
- Katz B, Schmitt OH (February 1940). "Electric interaction between two adjacent nerve fibres". J. Physiol. (Lond.). 97 (4): 471–88. doi:10.1113/jphysiol.1940.sp003823. PMC 1393925. PMID 16995178.
- Arvanitaki A (1942). "Effects Evoked in an Axon by the Activity of a Contiguous One". Journal of Neurophysiology. 5 (2): 89–108. doi:10.1152/jn.1942.5.2.89. ISSN 0022-3077.
- Jefferys JG (October 1995). "Nonsynaptic modulation of neuronal activity in the brain: electric currents and extracellular ions". Physiol. Rev. 75 (4): 689–723. doi:10.1152/physrev.1995.75.4.689. PMID 7480159.
- Ramón F, Moore JW (1978). "Ephaptic transmission in squid giant axons". Am. J. Physiol. 234 (5): C162–9. doi:10.1152/ajpcell.1978.234.5.C162. PMID 206154.
- Van der Goes van Naters W (2013). "Inhibition among olfactory receptor neurons". Front Hum Neurosci. 7: 690. doi:10.3389/fnhum.2013.00690. PMC 3805947. PMID 24167484.
- Anastassiou CA, Perin R, Markram H, Koch C (February 2011). "Ephaptic coupling of cortical neurons". Nat. Neurosci. 14 (2): 217–23. doi:10.1038/nn.2727. PMID 21240273. S2CID 7836328. (direct link to full text)
- Radman T, Su Y, An JH, Parra LC, Bikson M (March 2007). "Spike timing amplifies the effect of electric fields on neurons: implications for endogenous field effects". J. Neurosci. 27 (11): 3030–6. doi:10.1523/JNEUROSCI.0095-07.2007. PMC 6672570. PMID 17360926.
- Anastassiou CA, Montgomery SM, Barahona M, Buzsáki G, Koch C (February 2010). "The effect of spatially inhomogeneous extracellular electric fields on neurons". J. Neurosci. 30 (5): 1925–36. doi:10.1523/JNEUROSCI.3635-09.2010. PMC 6633973. PMID 20130201.
- Hrg D (April 2013). "Synchronization of two Hindmarsh-Rose neurons with unidirectional coupling". Neural Netw. 40: 73–9. doi:10.1016/j.neunet.2012.12.010. PMID 23454921.
- Pinotsis, Dimitris A; Miller, Earl K (2023-07-07). "In vivo ephaptic coupling allows memory network formation". Cerebral Cortex. doi:10.1093/cercor/bhad251. ISSN 1047-3211.
- Pinotsis, Dimitris A.; Fridman, Gene; Miller, Earl K. (2023-07-01). "Cytoelectric coupling: Electric fields sculpt neural activity and "tune" the brain's infrastructure". Progress in Neurobiology. 226: 102465. doi:10.1016/j.pneurobio.2023.102465. ISSN 0301-0082.
- Martinez-Banaclocha, Marcos (13 February 2020). "Astroglial Isopotentiality and Calcium-Associated Biomagnetic Field Effects on Cortical Neuronal Coupling". Cells. 9 (2): 439. doi:10.3390/cells9020439. PMC 7073214.
- Veeraraghavan R, Gourdie RG, Poelzing S (March 2014). "Mechanisms of cardiac conduction: a history of revisions". Am. J. Physiol. Heart Circ. Physiol. 306 (5): H619–27. doi:10.1152/ajpheart.00760.2013. PMC 3949060. PMID 24414064.
- Ivanovic, Ena; Kucera, Jan P. (November 2021). "Localization of Na + channel clusters in narrowed perinexi of gap junctions enhances cardiac impulse transmission via ephaptic coupling: a model study". The Journal of Physiology. 599 (21): 4779–4811. doi:10.1113/JP282105.
- Lin J, Keener JP (2010). "Modeling electrical activity of myocardial cells incorporating the effects of ephaptic coupling". Proc. Natl. Acad. Sci. U.S.A. 107 (49): 20935–40. Bibcode:2010PNAS..10720935L. doi:10.1073/pnas.1010154107. PMC 3000303. PMID 21078961.
- Veeraraghavan R, Hoeker GS, Alvarez-Laviada A, Hoagland D, Wan X, King DR, Sanchez-Alonso J, Chen C, Jourdan J, Isom LL, Deschenes I, Smyth JW, Gorelik J, Poelzing S, Gourdie RG (2018). "The adhesion function of the sodium channel beta subunit (β1) contributes to cardiac action potential propagation". eLife. 7. doi:10.7554/eLife.37610. PMC 6122953. PMID 30106376.
- Ivanovic, Ena; Kucera, Jan P. (2 November 2022). "Tortuous Cardiac Intercalated Discs Modulate Ephaptic Coupling". Cells. 11 (21): 3477. doi:10.3390/cells11213477.
- Hoagland, Daniel T.; Santos, Webster; Poelzing, Steven; Gourdie, Robert G. (July 2019). "The role of the gap junction perinexus in cardiac conduction: Potential as a novel anti-arrhythmic drug target". Progress in Biophysics and Molecular Biology. 144: 41–50. doi:10.1016/j.pbiomolbio.2018.08.003. hdl:10919/99879.
- Dudek FE, Yasumura T, Rash JE (November 1998). "'Non-synaptic' mechanisms in seizures and epileptogenesis". Cell Biol. Int. 22 (11–12): 793–805. doi:10.1006/cbir.1999.0397. PMID 10873292. S2CID 2092166.
- Stacey RG, Hilbert L, Quail T (May 2015). "Computational study of synchrony in fields and microclusters of ephaptically coupled neurons". J. Neurophysiol. 113 (9): 3229–41. doi:10.1152/jn.00546.2014. PMC 4440237. PMID 25673735.
- Bokil H, Laaris N, Blinder K, Ennis M, Keller A (October 2001). "Ephaptic interactions in the mammalian olfactory system". J. Neurosci. 21 (20): RC173. doi:10.1523/JNEUROSCI.21-20-j0004.2001. PMC 6763860. PMID 11588203.
- Faber, Donald S.; Funch, Paul G. (May 1980). "Differential properties of orthodromic and antidromic impulse propagation across the Mauthner cell initial segment". Brain Research. 190 (1): 255–260. doi:10.1016/0006-8993(80)91176-2. PMID 7378738. S2CID 10722904.
- Martin, A. R.; Pilar, G. (1964-07-01). "An analysis of electrical coupling at synapses in the avian ciliary ganglion". The Journal of Physiology. 171 (3): 454–475. doi:10.1113/jphysiol.1964.sp007390. PMC 1368844. PMID 14193934.
- Blot, Antonin; Barbour, Boris (February 2014). "Ultra-rapid axon-axon ephaptic inhibition of cerebellar Purkinje cells by the pinceau". Nature Neuroscience. 17 (2): 289–295. doi:10.1038/nn.3624. ISSN 1097-6256. PMID 24413696. S2CID 2515949.
- Sierksma, Martijn C.; Borst, J. Gerard G. (2021-10-26). Berry, Hugues (ed.). "Using ephaptic coupling to estimate the synaptic cleft resistivity of the calyx of Held synapse". PLOS Computational Biology. 17 (10): e1009527. doi:10.1371/journal.pcbi.1009527. ISSN 1553-7358. PMC 8570497. PMID 34699519.
- Bédard C, Destexhe A (August 2013). "Generalized cable theory for neurons in complex and heterogeneous media". Phys Rev E. 88 (2): 022709. arXiv:1304.5674. Bibcode:2013PhRvE..88b2709B. doi:10.1103/PhysRevE.88.022709. PMID 24032866. S2CID 16729315.
- Hand PE, Griffith BE (August 2010). "Adaptive multiscale model for simulating cardiac conduction". Proc. Natl. Acad. Sci. U.S.A. 107 (33): 14603–8. Bibcode:2010PNAS..10714603H. doi:10.1073/pnas.1008443107. PMC 2930419. PMID 20671202.
- Mori Y, Fishman GI, Peskin CS (April 2008). "Ephaptic conduction in a cardiac strand model with 3D electrodiffusion". Proc. Natl. Acad. Sci. U.S.A. 105 (17): 6463–8. Bibcode:2008PNAS..105.6463M. doi:10.1073/pnas.0801089105. PMC 2359793. PMID 18434544.
- Ren J, Momose-Sato Y, Sato K, Greer JJ (January 2006). "Rhythmic neuronal discharge in the medulla and spinal cord of fetal rats in the absence of synaptic transmission". J. Neurophysiol. 95 (1): 527–34. doi:10.1152/jn.00735.2005. PMID 16148265.
- Faber DS, Korn H (July 1989). "Electrical field effects: their relevance in central neural networks". Physiol. Rev. 69 (3): 821–63. doi:10.1152/physrev.1989.69.3.821. PMID 2546168.
- Roth BJ (February 2014). "Does ephaptic coupling contribute to propagation in cardiac tissue?". Biophys. J. 106 (4): 774–5. Bibcode:2014BpJ...106..774R. doi:10.1016/j.bpj.2014.01.011. PMC 3944891. PMID 24559978.
- Chiang, Chia-Chu; Shivacharan, Rajat S.; Wei, Xile; Gonzalez-Reyes, Luis E.; Durand, Dominique M. (2019). "Slow periodic activity in the longitudinal hippocampal slice can self-propagate non-synaptically by a mechanism consistent with ephaptic coupling". The Journal of Physiology. 597 (1): 249–269. doi:10.1113/JP276904. PMC 6312416. PMID 30295923.