Forelimb
A forelimb or front limb is one of the paired articulated appendages (limbs) attached on the cranial (anterior) end of a terrestrial tetrapod vertebrate's torso. With reference to quadrupeds, the term foreleg or front leg is often used instead. In bipedal animals with an upright posture (e.g. humans and some primates), the term upper limb is often used.
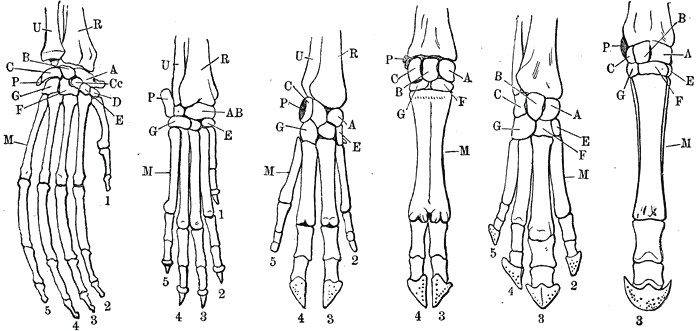
A forelimb is not to be confused with a forearm, which is a distal portion of the human upper limb between the elbow and the wrist.
All vertebrate forelimbs are homologous, meaning that they all evolved from the same structures. For example, the flipper of a turtle or of a dolphin, the arm of a human, the foreleg of a horse, and the wings of both bats and birds are ultimately homologous, despite the large differences between them.[1]
Specific uses of the forelimbs may be analogous if they evolved from different sub-structures of the forelimb, such as the flippers of turtles and dolphins, and the wings of birds and bats.[2]
Evolution of forelimbs
Evolution of the forelimb may be characterized by many trends. The number of digits, their characteristics, as well as the shape and alignment of radius, ulna, and humerus, have had major evolutionary implications.
Changes in body size, foot posture, habitat, and substrate are frequently found to influence one another (and to connect to broader potential drivers, such as changing climate).[3]
Shape
A number of factors can influence the evolution of forelimb long bone shape, such as body mass, lifestyle, predatory behavior, or relative prey size. A general pattern is for heavier species to have more robust radii, ulnas, and humeri.[4]
Musteloid carnivorans that have an arboreal lifestyle tend to have long and slender forelimb long bones, which allow for improved movement and flexibility. Semi-fossorial and aquatic musteloid species tend to have short and robust forelimb long bones to deal with the strain from digging and swimming.[5]
In the order Carnivora, felids, which usually ambush and grapple with their prey, have shorter and more robust limbs. Their forelimbs are used for both short sprints and grappling, which means that they need to be flexible and durable. In contrast, canids, which often pursue their prey over greater distances, have longer, more gracile limbs. Running is pretty much the only use for their forelimbs, so they do not need to be adapted for anything else and can be less flexible.[6]
Predators hunting prey that is half their body weight or greater evolved shorter and more sturdy radii, ulnas, and humeri to decrease the likelihood of the bone breaking or fracturing while hunting. Predators hunting prey less than half their body weight tended to have longer and more slender forelimb long bones to improve energetic efficiency.[7]
Polydactyly
Tetrapods were initially understood to have first developed five digits as an ancestral characteristic, which were then reduced or specialized into a number of uses. Certain animals retained 'primitive' forelimbs, such as pentadactylous (five-fingered) reptiles and primates. This has mostly held true, but the earliest tetrapod or "fishapod" ancestors may have had more than five digits. This was notably challenged by Stephen Jay Gould in his 1991 essay "Eight (Or Fewer) Little Piggies".[8]
Polydactyly in early tetrapods should be understood as having more than five digits to the finger or foot, a condition that was the natural state of affairs in the very first tetrapods. Early groups like Acanthostega had eight digits, while the more derived Ichthyostega had seven digits, the yet-more derived Tulerpeton had six toes.
Tetrapods evolved from animals with fins such as found in lobe-finned fishes. From this condition a new pattern of limb formation evolved, where the development axis of the limb rotated to sprout secondary axes along the lower margin, giving rise to a variable number of very stout skeletal supports for a paddle-like foot.
Digit specialization
Digits may be specialized for different forms of locomotion. A classic example is the horse's development of a single toe (monodactyly).[3] Other hooves, like those of even-toed and odd-toed ungulates, and even the hoof-like foot of extinct hadrosaurs,[9] may be regarded as similar specializations.
To bear their immense weight, sauropods, the most derived being titanosaurs, developed a tubular manus (front foot) and gradually lost their digits, standing on their metacarpals.[10] The stegosaurian forelimb has evidence for a sauropod−like metacarpal configuration[11] This was a different evolutionary strategy than megafaunal mammals such as modern elephants.
Therapsids started evolving diverse and specialized forelimbs 270 million years ago, during the Permian.[12]
Opposable thumbs
Modern humans are unique in the musculature of the forearm and hand, though opposable thumbs or structures like them have arisen in a few animals.
In dinosaurs, a primitive autonomization of the first carpometacarpal joint (CMC) may have occurred. In primates, a real differentiation appeared perhaps 70 mya, while the shape of the human thumb CMC finally appears about 5 mya.
- Primates fall into one of four groups:[13]
- Nonopposable thumbs: tarsiers and marmosets
- Pseudo-opposable thumbs: all strepsirrhines and Cebidae
- Opposable thumbs: Old World monkeys and all great apes
- Opposable with comparatively long thumbs: gibbons (or lesser apes)
Pandas have evolved pseudo-opposable thumbs by extension of the sesamoid bone, which is not a true digit.[14]
Pronation and supination
The ability to pronate the manus (hand) and forearm in therian mammals is achieved by a rounded head of the radius, which allows it to swivel across the ulna. Supination requires a dorsal glide of the distal radius and pronation a palmar glide in relation to the distal ulna.
Pronation has evolved multiple times, among mammals, chameleons, and varanids.[15] However, the more basal condition is to be unable to pronate. Dinosaurs were not capable of more than semi-pronation of the wrist,[16] though bipedal origins of all quadrupedal dinosaur clades could have allowed for greater disparity in forelimb posture than often considered.[15] Monotremes have forearms that are not as dexterous as therians. Monotremes have a sprawling posture, and multiple elements in their pectoral girdles, which are ancestral traits for mammals.[17]
In birds, the forearm muscles supinate, pronate, flex and extend the distal wing.[18]
Wings
All tetrapod forelimbs are homologous, evolving from the same initial structures in lobe-finned fish. However, another distinct process may be identified, convergent evolution, by which the wings of birds, bats, and extinct pterosaurs evolved the same purpose in drastically different ways.[2] These structures have similar form or function but were not present in the last common ancestor of those groups.
Bat wings are composed largely of a thin membrane of skin supported on the five fingers, whereas bird wings are composed largely of feathers supported on much reduced fingers, with finger 2 supporting the alula and finger 4 the primary feathers of the wing; there are only distant homologies between birds and bats, with much closer homologies between any pair of bird species, or any pair of bat species.
Flippers
Marine mammals have evolved several times. Over the course of their evolution, they develop streamlined hydrodynamic bodies. The forelimb thus develops into a flipper. The forelimbs of cetaceans, pinnipeds, and sirenians presents a classic example of convergent evolution. There is widespread convergence at the gene level.[19] Distinct substitutions in common genes created various aquatic adaptations, most of which constitute parallel evolution because the substitutions in question are not unique to those animals.[20]
When comparing cetaceans to pinnipeds to sirenians, 133 parallel amino acid substitutions occur. Comparing and contrasting cetaceans-pinnipeds, cetaceans-sirenians, and pinnipeds-sirenians, 2,351, 7,684, and 2,579 substitutions occur, respectively.[20]
See also
Bibliography
- de Beer, Gavin (1956). Vertebrate zoology: an introduction to the comparative anatomy, embryology, and evolution of chordate animals. Sidgwick and Jackson.
References
- "Image Gallery: Homo Sapiens. homology: homologies of the forelimb among vertebrates". Encyclopædia Britannica. Retrieved January 27, 2013.
- "Homologies and analogies". evolution.berkeley.edu. Retrieved 2019-12-09.
- McHorse, Brianna K.; Biewener, Andrew A.; Pierce, Stephanie E. (2019-09-01). "The Evolution of a Single Toe in Horses: Causes, Consequences, and the Way Forward". Integrative and Comparative Biology. 59 (3): 638–655. doi:10.1093/icb/icz050. ISSN 1540-7063. PMID 31127281.
- Fabre, Anne-Claire; Cornette, Raphael; Peigné, Stéphane; Goswami, Anjali (2013-05-21). "Influence of body mass on the shape of forelimb in musteloid carnivorans". Biological Journal of the Linnean Society. 110 (1): 91–103. doi:10.1111/bij.12103. ISSN 0024-4066.
- Fabre, Anne-Claire; Cornette, Raphael; Goswami, Anjali; Peigné, Stéphane (2015-05-21). "Do constraints associated with the locomotor habitat drive the evolution of forelimb shape? A case study in musteloid carnivorans". Journal of Anatomy. 226 (6): 596–610. doi:10.1111/joa.12315. ISSN 0021-8782. PMC 4450962. PMID 25994128.
- Meachen-Samuels, Julie; Van Valkenburgh, Blaire (June 2009). "Forelimb indicators of prey-size preference in the Felidae". Journal of Morphology. 270 (6): 729–744. doi:10.1002/jmor.10712. ISSN 0362-2525. PMID 19123240. S2CID 20732642.
- Michaud, Margot; Veron, Géraldine; Fabre, Anne‐Claire (2020-11-06). "Phenotypic integration in feliform carnivores: Covariation patterns and disparity in hypercarnivores versus generalists". Evolution. 74 (12): 2681–2702. doi:10.1111/evo.14112. ISSN 0014-3820. PMID 33085081. S2CID 224824184.
- Stephen Jay Gould. "Stephen Jay Gould "Eight (or Fewer) Little Piggies" 1991". Archived from the original on 2010-01-11. Retrieved 2015-10-02.
- Zheng, R. ; Farke (2011). "A Photographic Atlas of the Pes from a Hadrosaurine Hadrosaurid Dinosaur". PalArch's Journal of Vertebrate Palaeontology. 8 (7): 1–12. ISSN 1567-2158.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Apesteguía, Sebastián (2005-01-01). "Evolution of the titanosaur metacarpus". Thunder-Lizards: The Sauropodomorph Dinosaurs: 321–345.
- Senter, Phil (2010). "Evidence for a Sauropod-Like Metacarpal Configuration in Stegosaurian Dinosaurs". Acta Palaeontologica Polonica. 55 (3): 427–432. doi:10.4202/app.2009.1105. ISSN 0567-7920.
- "Mammals' unique arms started evolving before the dinosaurs existed". ScienceDaily. Retrieved 2019-12-10.
- Ankel-Simons, Friderun. (2007). Primate anatomy : an introduction (3rd ed.). Amsterdam: Elsevier Academic Press. ISBN 978-0-08-046911-9. OCLC 437597677.
- Salesa, Manuel J.; Antón, Mauricio; Peigné, Stéphane; Morales, Jorge (2006-01-10). "Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas". Proceedings of the National Academy of Sciences. 103 (2): 379–382. Bibcode:2006PNAS..103..379S. doi:10.1073/pnas.0504899102. ISSN 0027-8424. PMC 1326154. PMID 16387860.
- VanBuren, Collin S.; Bonnan, Matthew (2013-09-18). "Forearm Posture and Mobility in Quadrupedal Dinosaurs". PLOS ONE. 8 (9): e74842. Bibcode:2013PLoSO...874842V. doi:10.1371/journal.pone.0074842. ISSN 1932-6203. PMC 3776758. PMID 24058633.
- Hutson, Joel D. (2014). "Quadrupedal Dinosaurs did not Evolve Fully Pronated Forearms: New Evidence from the Ulna". Acta Palaeontologica Polonica. 60 (3): 599–610. doi:10.4202/app.00063.2014. ISSN 0567-7920.
- Hall, Brian Keith, 1941- (2007). Fins into limbs evolution, development, and transformation. University of Chicago Press. OCLC 928978489.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Tobalske, Bret W. (2007-09-15). "Biomechanics of bird flight". Journal of Experimental Biology. 210 (18): 3135–3146. doi:10.1242/jeb.000273. ISSN 0022-0949. PMID 17766290.
- Chikina, Maria; Robinson, Joseph D.; Clark, Nathan L. (2016-09-01). "Hundreds of Genes Experienced Convergent Shifts in Selective Pressure in Marine Mammals". Molecular Biology and Evolution. 33 (9): 2182–2192. doi:10.1093/molbev/msw112. ISSN 0737-4038. PMC 5854031. PMID 27329977.
- Zhou, Xuming; Seim, Inge; Gladyshev, Vadim N. (2015-11-09). "Convergent evolution of marine mammals is associated with distinct substitutions in common genes". Scientific Reports. 5 (1): 16550. Bibcode:2015NatSR...516550Z. doi:10.1038/srep16550. ISSN 2045-2322. PMC 4637874. PMID 26549748.
