Haplogroup B-M60
Haplogroup B (M60) is a human Y-chromosome DNA haplogroup common to paternal lineages in Africa. It is a primary branch of the haplogroup BT.
| Haplogroup B | |
|---|---|
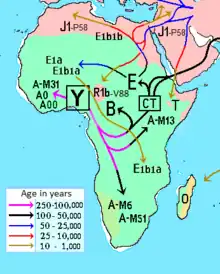 The geographical concentrations of major haplogroups, including B (M60), in Africa. | |
| Possible time of origin | 100,000 years BP[1] |
| Possible place of origin | Africa,[2] region unknown |
| Ancestor | BT |
| Descendants | Primary: B1 (M236), B2 (M182), B3 (L1387); Subclades of the above include: B1a (M146); B2b (M112); B2a1a1a1 (M109) |
| Defining mutations | M60, M181/Page32, P85, P90, V62, V75, V78, V83, V84, V85, V90, V93, V94, V185, V197, V217, V227, V234, V237, and V44 |
| Highest frequencies | Baka 63% (Gabon & Cameroon)[3] - 72% (CAR),[4] Hadzabe (Tanzania) 52%[5]-60%,[6] Nuer (South Sudan) 50%,[7] Mbuti (DRC) 33%[8]-60%,[4] Biaka (CAR) 35%[8]-55%,[4] Central Africa 32%,[9] Tsumkwe San (Namibia) 31%,[4] Khoisan 28%,[9] Shilluk (South Sudan) 27%,[7] Burunge (Tanzania) 25%,[6] Dinka (South Sudan) 23%,[7] Ngumba (Cameroon) 23%[4]-33%,[3] Eviya (Gabon) 21%,[3] Fali (Cameroon) 18%,[8] Sotho–Tswana (South Africa) 18%,[4] Zulu (South Africa) 17%,[4] Eshira (Gabon) 17%,[3] Shake (Gabon) 16%,[3] Hausa (Sudan) 16%,[7] Sukuma (Tanzania) 16%,[5] Bakola (Cameroon) 15%[4]-36%,[3] Copts (Sudan) 15%,[7] Sudan 15%,[9] Kunama (Eritrea) 15%,[10] Tutsi (Rwanda) 15%,[11] Sandawe (Tanzania) 15%,[6] Uldeme (Cameroon) 5%[8]-31%,[4] Nuba (Sudan) 14%,[7] Makina (Gabon) 14%,[3] Southern Africa 13%,[9] Mali 11%,[9] Ewondo (Cameroon) 10%,[8] Ethiopia 10%,[9] Shona (Zimbabwe) 10%[4] Qeshmi (Iran) 8,2%,[12] Bandari (Iran) 2,3%,[12] Hazara (Afghanistan) 5,1%,[13] |
B (M60) is common in parts of Africa, especially the tropical forests of West-Central Africa. It was the ancestral haplogroup of not only modern Pygmies like the Baka and Mbuti, but also Hadzabe from Tanzania, who often have been considered, in large part because of some typological features of their language, to be a remnant of Khoisan people in East Africa.
Distribution
According to one study of the Y-DNA of populations in Sudan, haplogroup B-M60 is found in approximately 30% (16/53) of Southern Sudanese, 16% (5/32) of local Hausa people, 14% (4/28) of the Nuba of central Sudan, 3.7% (8/216) of Northern Sudanese (but only among Copts and Nubians), and 2.2% (2/90) of Western Sudanese.[7] According to another study, haplogroup B is found in approximately 15% of Sudanese males, including 12.5% (5/40) B2a1a1a1 (M109/M152) and 2.5% (1/40) B-M60(xM146, M150, M112).[9]
In Madagascar, haplogroup B-M60 has been found in approximately 9% of Malagasy males, including 6% (2/35) B-M60(xB2b-50f2(P)) and 3% (1/35) B2b-50f2(P).[14]
Family Tree DNA shows a significant number of persons of Haplogroup B-M60 (B-M181) claiming origins from the Arabian Peninsula (dominantly Saudi Arabia, but also in Kuwait, Bahrain, Yemen, Qatar, Iraq, United Arab Emirates, and Oman).[15] Sampling bias does not allow for meaningful percentages, but the presence of the haplogroup is solidly attested.
In Hormozgan Province in Iran, haplogroup B-M60 has been found in 8.2% of a sample of 49 Qeshmi people, and in 2.3% of a sample of 131 Bandari people.[16]
In Afghanistan, haplogroup B-M60 has been found in 5.1% (3/59) of a sample of Hazara males.[17]
In United Kingdom, haplogroup B-M60(xM218) has been found by FTDNA in 1 individual.
Subclades
B-M236
Haplogroup B-M236 has been found in 4% (2/48) of a sample of Bamileke males from southern Cameroon.[8]
B-M146
Haplogroup B-M146 has been found in 2% (1/49) of a sample of Mossi males from Burkina Faso[8] and in 2% (1/44) of a sample of unspecified ethnic affiliation from Mali.[9]
B-M182
Haplogroup B-M182 has been found in 6% (3/47) of a sample of Mbuti males from the Democratic Republic of the Congo, 6% (2/33) of a sample of Bakola males from southern Cameroon,[4] 6% (1/18) of a sample of Dama males from Namibia,[4] and 3% (1/31) of a sample of Biaka males from Central African Republic.[4] The vast majority of Family Tree DNA participants in Haplogroup B-M60 test positive for B-M182, with three-fourths of those participants claiming countries of the Arabian Peninsula as their ancestral land of origin,[15] attesting to its presence in that area also.
B-M150
Haplogroup B-M150 has been found in 8% (1/12) of a sample of Mbuti males from the Democratic Republic of the Congo.[8]
Haplogroup B-M150(xM152) has been observed in 11% (5/47) of a sample of Mbuti from Democratic Republic of the Congo, 11% (1/9) of a sample of Tupuri from northern Cameroon, 11% (1/9) of a sample of Luo from Kenya, 7% (4/55) of a sample of Dogon from Mali, 6% (1/18) of a sample of Baka from Central African Republic, and 2% (1/42) of a sample of Kikuyu and Kamba from Kenya.[4]
Haplogroup B-M150(xM109/M152, M108.1) has been found in 3% (1/37) of a sample from Central Africa, 2% (1/44) of a sample from Mali, and 1% (1/88) of a sample from Ethiopia.[9]
Without testing for any downstream mutation, haplogroup B-M150 has been found in 33.3% (8/24) of a sample of Ngumba from Cameroon,[3] 20.8% (5/24) of a sample of Eviya from Gabon,[3] 18.2% (4/22) of a sample of Bakola from Cameroon,[3] 14.3% (6/42) of a sample of Eshira from Gabon,[3] 14.0% (6/43) of a sample of Makina from Gabon,[3] 14.0% (6/43) of a sample of Shake from Gabon,[3] 8.6% (5/58) of a sample of Punu from Gabon,[3] 8.3% (5/60) of a sample of Tsogo from Gabon,[3] 7.0% (4/57) of a sample of Nzebi from Gabon,[3] 6.7% (1/15) of a sample of Mbugwe from Tanzania,[6] 4.3% (2/46) of a sample of Duma from Gabon,[3] 4.3% (2/47) of a sample of Obamba from Gabon,[3] 4.2% (2/48) of a sample of Benga from Gabon,[3] 3.8% (2/53) of a sample of Kota from Gabon,[3] 2.8% (1/36) of a sample of Ndumu from Gabon,[3] 2.1% (1/47) of a sample of Galoa from Gabon,[3] 2.0% (1/50) of a sample of Akele from Gabon,[3] 1.7% (1/60) of a sample of Fang from Gabon,[3] 1.5% (1/68) of a sample of Sandawe from Tanzania,[6] 1.4% (1/72) of a sample from Qatar,[18] and 0.64% (1/157) of a sample from Saudi Arabia.[19]
B-M218
Haplogroup B-M218 has been found in 17% (20/118) of a mixed sample of Nilotic ethnic groups of Karamojong, Jie and Dodos from Karamoja region in Uganda.[20] This haplogroup has also been found by FTDNA in 1 individual from Qatar, 3 individuals from Saudi Arabia,[21] 1 individual from Syria, 1 individual from Tunisia, 1 individual from United Kingdom.
B-M109
Haplogroup B2a1a1a1 (M109, M152, P32), previously B2a1a is the most commonly observed subclade of haplogroup B.
In Central Africa, B-M109 Y-DNA has been found in 23% (7/31) of Ngumba males from southern Cameroon,[4] 18% (7/39) of Fali males from northern Cameroon,[8] 5% (1/21)[8] to 31% (4/13)[4] of Uldeme males from northern Cameroon, 10% (3/29) of Ewondo males from southern Cameroon,[8] 7% (1/15) of a mixed sample of speakers of various Chadic languages from northern Cameroon,[8] 6% (1/18) of a mixed sample of speakers of various Adamawa languages from northern Cameroon,[8] 6% (2/33) of Bakola males from southern Cameroon,[4] 4% (1/28) of Mandara males from northern Cameroon,[4] and 3% (1/31)[4] to 5% (1/20)[8] of Biaka males from Central African Republic.
In East Africa, haplogroup B2a1a1a1 Y-DNA has been found in 11% (1/9) of a small sample of Iraqw males from Tanzania,[4] 11% (1/9) of a small sample of Luo males from Kenya,[4] 8% (2/26) of Maasai males from Kenya,[4] and 4.5% (4/88) of a sample of Ethiopians.[9]
In Southern Africa, B-M109 Y-DNA has been found in 18% (5/28) of Sotho–Tswana males from South Africa,[4] 14% (4/29) of Zulu males from South Africa,[4] 13% (7/53) of an ethnically mixed sample of non-Khoisan Southern Africans,[9] 10% (5/49) of Shona males from Zimbabwe,[4] and 5% (4/80) of Xhosa males from South Africa.[4]
In North Africa, haplogroup B2a1a1a1 Y-DNA has been found in 12.5% (5/40) of Sudanese[9] and 2% (2/92) of Egyptians.[4]
In Eurasia, B2a1a1a1 (B-M109) has been found in 3% (3/117) of a sample of Iranians from southern Iran[22] and 2% (2/88) of a sample from Pakistan and India.[9]
B-M108.1
Haplogroup B-M108.1 (M108.1) has been found in 3% (3/88) of a sample from Ethiopia.[9]
B-M43
Haplogroup B-M43 (M43, P111) has been found in 7% (3/44) of a sample from Mali.[9]
B-M112
Haplogroup B-M112 (M112, M192, 50f2(P)) has been found mainly among pygmy populations in Central Africa, Juu (Northern Khoisan) populations in Southern Africa, and the Hadzabe in East Africa. It also has been found occasionally in samples of groups who neighbor the aforementioned populations.
Specifically, haplogroup B2b has been observed in 67% (12/18) of a sample of Baka from Central African Republic,[4] 52% (12/23) or 51% (29/57) of a sample of Hadzabe from Tanzania,[5][6] 48% (15/31) of a sample of Biaka from Central African Republic,[4] 43% (20/47) of a sample of Mbuti from the Democratic Republic of the Congo,[4] 31% (9/29) of a sample of Tsumkwe San from Namibia,[4] 28% (11/39) of a sample of the Northern Khoisan-speaking Ju|’hoansi and Sekele peoples,[5][9] 25% (6/24) of a sample of Burunge from Tanzania,[6] 14% (13/94) of a sample of Tutsi from Rwanda,[11] 13% (9/68) of a sample of Sandawe from Tanzania,[6] 9% (3/32) of a sample of !Kung/Sekele from Namibia,[4] 5% (1/20) of a sample of Turu from Tanzania,[6] 5% (2/43) of a sample of Wairak from Tanzania,[11] 3% (1/29) of a sample of Zulu from South Africa,[4] 3% (1/33) of a sample of Bakola from southern Cameroon,[4] 3% (1/35) of a sample of Datog from Tanzania,[6] 3% (1/35) of a sample of Malagasy,[14] 1.4% (1/69) of a sample of Hutu from Rwanda,[11] 1.4% (1/72) of a sample from Qatar,[18] and 1.3% (2/157) of a sample from Saudi Arabia.[19]
B-P6
Haplogroup B-P6 has been found in Khoisan populations of Namibia, including 24% (7/29) of a sample of Tsumkwe San and 3% (1/32) of a sample of !Kung/Sekele.[4]
B-M115
Haplogroup B-M115 has been found in 8% (1/12) of a sample of Mbuti from the Democratic Republic of the Congo.[8]
B-M30
Haplogroup B-M30 has been found in 22% (2/9) of a mixed sample of speakers of Central Sudanic and Saharan languages from northern Cameroon and in 5% (1/20) of a sample of Biaka from Central African Republic.[8]
B-M108.2
Haplogroup B-M108.2 has been found in 25% (1/4) of a very small sample of Lissongo from Central African Republic.[8]
B-P7
Haplogroup B-P7 has been observed most frequently in samples of some populations of pygmies from Central Africa: 67% (12/18) Baka from Central African Republic,[4] 45% (14/31) Biaka from Central African Republic,[4] 21% (10/47) Mbuti from Democratic Republic of the Congo.[4] This haplogroup also has been found in an Iraqw (South Cushitic) individual from Tanzania (1/9 = 11%) and in some samples of Khoisan from Namibia (2/32 = 6% !Kung/Sekele, 2/29 = 7% Tsumkwe San).[4]
Phylogenetics
Phylogenetic history
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome Phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Longhand) | YCC 2005 (Longhand) | YCC 2008 (Longhand) | YCC 2010r (Longhand) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B-M60 | 2 | II | 1B | 5 | - | H1 | B | B* | B | B | B | B | B | B | B | B | B | B |
| B-M146 | 2 | II | 1B | 5 | - | H1 | B | B1 | B1 | B1a | B1a | B1a | B1a | B1a | B1a | B1a | B1a | B1a |
| B-M182 | * | * | * | * | * | * | * | * | B2 | B2 | B2 | B2 | B2 | B2 | B2 | B2 | B2 | B2 |
| B-M150 | 2 | II | 1B | 5 | - | H1 | B | B2a* | B2a | B2a | B2a | B2a | B2a | B2a | B2a | B2a | B2a | B2a |
| B-M109 | 2 | II | 1B | 5 | - | H1 | B | B2a1 | B2a1 | B2a1a | B2a1a | B2a1a | B2a1a | B2a1a | B2a1a | B2a1a | B2a1a | B2a1a |
| B-M108.1 | 2 | II | 1B | 5 | - | H1 | B | B2a2* | B2a2 | B2a2 | B2a2 | B2a2 | B2a2 | B2a2 | B2a2 | B2a2 | removed | removed |
| B-M43 | 2 | II | 1B | 5 | - | H1 | B | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a | B2a2a |
| B-M112 | 6 | II | 1B | 6 | - | H1 | B | B2b* | B2b | B2b | B2b | B2b | B2b | B2b | B2b | B2b | B2b | B2b |
| B-P6 | 6 | II | 1B | 7 | - | H1 | B | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 | B2b1 |
| B-M115 | 6 | II | 1B | 6 | - | H1 | B | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 | B2b2 |
| B-M30 | 6 | II | 1B | 6 | - | H1 | B | B2b3* | B2b3 | B2b3 | B2b3 | B2b3 | B2b3 | B2b3 | B2b3 | B2b3 | B2b3 | B2b3 |
| B-M108.2 | 6 | II | 1B | 6 | - | H1 | B | B2b3a | B2b3a | B2b3a | B2b3a | B2b3a | B2b3a | B2b3a | B2b3a | B2b3a | removed | removed |
| B-P7 | 6 | II | 1B | 8 | - | H1 | B | B2b4* | B2b4 | B2b4 | B2b4 | B2b4 | B2b4 | B2b4 | B2b4 | B2b4 | removed | removed |
| B-P8 | 6 | II | 1B | 10 | - | H1 | B | B2b4a | B2b4a | B2b4a | B2b4a | B2b4a | B2b4a | B2b4a | B2b4a | B2b4a | removed | removed |
| B-M211 | 6 | II | 1B | 9 | - | H1 | B | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b | B2b4b |
Original research publications
The following research teams per their publications were represented in the creation of the YCC Tree.
Phylogenetic trees
The phylogenetic tree of haplogroup B subclades is based on the YCC 2008 tree[24] and subsequent published research.
- B
- B-M60 (M60, M181, P85, P90)
- B-M236 (M236, M288)
- B-M236 (M236)
- B-M182 (M182)
- B-M150 (M150)
- B-M218 (M218)
- B-M109 (M109, M152, P32)
- B-G1 (G1)
- B-M108.1 (M108.1)
- B-P111 (P111, M43)
- B-M218 (M218)
- B-M112 (M112, M192, 50f2(P))
- B-P6 (P6)
- B-M115 (M115, M169)
- B-M30 (M30, M129)
- B-M108.2 (M108.2)
- B-P7 (P7)
- B-P8 (P8, P70)
- B-MSY2.1 (MSY2.1, M211)
- B-P112 (P112)
- B-M150 (M150)
- B-M236 (M236, M288)
- B-M60 (M60, M181, P85, P90)
See also
Genetics
Y-DNA B subclades
- B-M108.1
- B-M108.2
- B-M109
- B-M112
- B-M115
- B-M146
- B-M150
- B-M182
- B-M211
- B-M30
- B-M43
- B-M60
- B-P6
- B-P7
- B-P8
Y-DNA backbone tree
References
- Kamin M, Saag L, Vincente M, et al. (April 2015). "A recent bottleneck of Y chromosome diversity coincides with a global change in culture". Genome Research. 25 (4): 459–466. doi:10.1101/gr.186684.114. PMC 4381518. PMID 25770088.
- Cruciani, Fulvio; Trombetta, Beniamino; Massaia, Andrea; Destro-Bisol, Giovanni; Sellitto, Daniele; Scozzari, Rosaria (2011). "A Revised Root for the Human y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa". The American Journal of Human Genetics. 88 (6): 814–818. doi:10.1016/j.ajhg.2011.05.002. PMC 3113241. PMID 21601174.
- Gemma Berniell-Lee, Francesc Calafell, Elena Bosch et al., "Genetic and demographic implications of the Bantu expansion: insights from human paternal lineages," Molecular Biology and Evolution Advance Access published April 15, 2009
- Elizabeth T Wood, Daryn A Stover, Christopher Ehret et al., "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes," European Journal of Human Genetics (2005) 13, 867–876. (cf. Appendix A: Y Chromosome Haplotype Frequencies)
- Knight, Alec; Underhill, Peter A.; Mortensen, Holly M.; et al. (March 2003). "African Y Chromosome and mtDNA Divergence Provides Insight into the History of Click Languages". Current Biology. 13 (6): 464–473. doi:10.1016/s0960-9822(03)00130-1. PMID 12646128. S2CID 52862939.
- Tishkoff, Sarah A.; Katherine Gonder, Mary; Henn, Brenna M.; et al. (2007). "History of Click-Speaking Populations of Africa Inferred from mtDNA and Y Chromosome Genetic Variation". Molecular Biology and Evolution. 24 (10): 2180–2195. doi:10.1093/molbev/msm155. PMID 17656633.
- Hassan, Hisham Y.; et al. (2008). "Y‐chromosome variation among Sudanese: Restricted gene flow, concordance with language, geography, and history". American Journal of Physical Anthropology. 137 (3): 316–323. doi:10.1002/ajpa.20876. PMID 18618658. Retrieved 11 October 2017.
- Cruciani, Fulvio; Santolamazza, Piero; Shen, Peidong; et al. (2002). "A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". American Journal of Human Genetics. 70 (5): 1197–1214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- Underhill PA, Shen P, Lin AA, et al. (November 2000). "Y chromosome sequence variation and the history of human populations". Nat. Genet. 26 (3): 358–61. doi:10.1038/81685. PMID 11062480. S2CID 12893406.
- Fulvio Cruciani, Beniamino Trombetta, Daniele Sellitto et al., "Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan connections and the spread of Chadic languages," European Journal of Human Genetics (2010), 1–8
- Luis, J. R.; Rowold, D. J.; Regueiro, M.; et al. (2004). "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations". American Journal of Human Genetics. 74 (3): 532–544. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- Viola Grugni, Vincenza Battaglia, Baharak Hooshiar Kashani, Silvia Parolo, Nadia Al-Zahery, et al. "Ancient Migratory Events in the Middle East : New Clues from the Y-Chromosome Variation of Modern Iranians" (2012)
- Haber M, Platt DE, Ashrafian Bonab M, Youhanna SC, Soria-Hernanz DF, et al. "Afghanistan's Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events" (2012)
- Hurles, Matthew E.; Sykes, Bryan C.; Jobling, Mark A.; Forster, Peter (2005). "The Dual Origin of the Malagasy in Island Southeast Asia and East Africa: Evidence from Maternal and Paternal Lineages". American Journal of Human Genetics. 76 (5): 894–901. doi:10.1086/430051. PMC 1199379. PMID 15793703.
- Family Tree DNA public haplotree, Haplogroup B-M181
- Viola Grugni, Vincenza Battaglia, Baharak Hooshiar Kashani, Silvia Parolo, Nadia Al-Zahery, et al. "Ancient Migratory Events in the Middle East : New Clues from the Y-Chromosome Variation of Modern Iranians", 2012
- Haber M, Platt DE, Ashrafian Bonab M, Youhanna SC, Soria-Hernanz DF, et al. "Afghanistan's Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events", 2012
- Cadenas, Alicia M; Zhivotovsky, Lev A; Cavalli-Sforza, Luca L; et al. (2008). "Y-chromosome diversity characterizes the Gulf of Oman". European Journal of Human Genetics. 16 (3): 374–386. doi:10.1038/sj.ejhg.5201934. PMID 17928816.
- Abu-Amero, Khaled K.; Hellani, Ali; Gonzalez, Ana M.; et al. (2009). "Saudi Arabian Y-Chromosome diversity and its relationship with nearby regions". BMC Genetics. 10: 59. doi:10.1186/1471-2156-10-59. PMC 2759955. PMID 19772609.
- Gomes, V; Sánchez-Diz, P; Amorim, A; Carracedo, A; Gusmão, L (March 2010). "Digging deeper into East African human Y chromosome lineages". Hum. Genet. 127 (5): 603–13. doi:10.1007/s00439-010-0808-5. PMID 20213473. S2CID 23503728.
- Middle East DNA Project
- Regueiro M.; Cadenas A.M.; Gayden T.; Underhill P.A.; Herrera R.J. (2006). "Iran: Tricontinental Nexus for Y-Chromosome Driven Migration". Human Heredity. 61 (3): 132–143. doi:10.1159/000093774. PMID 16770078. S2CID 7017701.
- Gomes, Verónica; Paula Sánchez-Diz; António Amorim; Ángel Carracedo; Leonor Gusmão (6 Mar 2010). "Digging deeper into East African human Y chromosome lineages". Human Genetics. 127 (5): 603–613. doi:10.1007/s00439-010-0808-5. PMID 20213473. S2CID 23503728.
- Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–8. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
Sources for conversion tables
- Capelli, Cristian; Wilson, James F.; Richards, Martin; Stumpf, Michael P.H.; et al. (February 2001). "A Predominantly Indigenous Paternal Heritage for the Austronesian-Speaking Peoples of Insular Southeast Asia and Oceania". The American Journal of Human Genetics. 68 (2): 432–443. doi:10.1086/318205.
- Hammer, Michael F.; Karafet, Tatiana M.; Redd, Alan J.; Jarjanazi, Hamdi; et al. (1 July 2001). "Hierarchical Patterns of Global Human Y-Chromosome Diversity". Molecular Biology and Evolution. 18 (7): 1189–1203. doi:10.1093/oxfordjournals.molbev.a003906.
- Jobling, Mark A.; Tyler-Smith, Chris (2000), "New uses for new haplotypes", Trends in Genetics, 16 (8): 356–62, doi:10.1016/S0168-9525(00)02057-6, PMID 10904265
- Kaladjieva, Luba; Calafell, Francesc; Jobling, Mark A; Angelicheva, Dora; et al. (February 2001). "Patterns of inter- and intra-group genetic diversity in the Vlax Roma as revealed by Y chromosome and mitochondrial DNA lineages". European Journal of Human Genetics. 9 (2): 97–104. doi:10.1038/sj.ejhg.5200597.
- Karafet, Tatiana; Xu, Liping; Du, Ruofu; Wang, William; et al. (September 2001). "Paternal Population History of East Asia: Sources, Patterns, and Microevolutionary Processes". The American Journal of Human Genetics. 69 (3): 615–628. doi:10.1086/323299.
- Semino, O.; Passarino, G; Oefner, PJ; Lin, AA; et al. (2000), "The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective", Science, 290 (5494): 1155–9, Bibcode:2000Sci...290.1155S, doi:10.1126/science.290.5494.1155, PMID 11073453
- Su, Bing; Xiao, Junhua; Underhill, Peter; Deka, Ranjan; et al. (December 1999). "Y-Chromosome Evidence for a Northward Migration of Modern Humans into Eastern Asia during the Last Ice Age". The American Journal of Human Genetics. 65 (6): 1718–1724. doi:10.1086/302680.
- Underhill, Peter A.; Shen, Peidong; Lin, Alice A.; Jin, Li; et al. (November 2000). "Y chromosome sequence variation and the history of human populations". Nature Genetics. 26 (3): 358–361. doi:10.1038/81685.