Histone-modifying enzymes
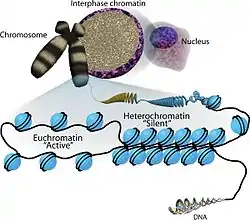
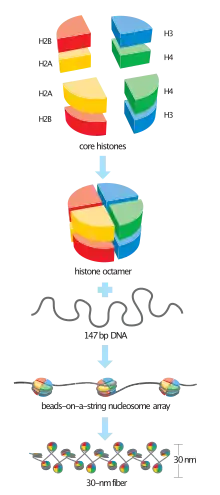
Histone-modifying enzymes are enzymes involved in the modification of histone substrates after protein translation and affect cellular processes including gene expression.[1][2] To safely store the eukaryotic genome, DNA is wrapped around four core histone proteins (H3, H4, H2A, H2B), which then join to form nucleosomes. These nucleosomes further fold together into highly condensed chromatin, which renders the organism's genetic material far less accessible to the factors required for gene transcription, DNA replication, recombination and repair.[3][4] Subsequently, eukaryotic organisms have developed intricate mechanisms to overcome this repressive barrier imposed by the chromatin through histone modification, a type of post-translational modification which typically involves covalently attaching certain groups to histone residues. Once added to the histone, these groups (directly or indirectly) elicit either a loose and open histone conformation, euchromatin, or a tight and closed histone conformation, heterochromatin. Euchromatin marks active transcription and gene expression, as the light packing of histones in this way allows entry for proteins involved in the transcription process. As such, the tightly packed heterochromatin marks the absence of current gene expression.[4]
While there exist several distinct post-translational modifications for histones, the four most common histone modifications include acetylation,[5] methylation,[6] phosphorylation[7] and ubiquitination.[8] Histone-modifying enzymes that induce a modification (e.g., add a functional group) are dubbed writers, while enzymes that revert modifications are dubbed erasers. Furthermore, there are many uncommon histone modifications including O-GlcNAcylation,[9] sumoylation,[10] ADP-ribosylation,[11] citrullination[12][13][14] and proline isomerization.[15] For a detailed example of histone modifications in transcription regulation see RNA polymerase control by chromatin structure and table "Examples of histone modifications in transcriptional regulation".
Common histone modifications
The four common histone modifications and their respective writer and eraser enzymes are shown in the table below.[16][17][18][19][20][21][22][23][24][25][26][27]
| Modification | Writer(s) | Eraser(s) | Effect on DNA |
|---|---|---|---|
| Acetylation | Histone acetyltransferases (HATs) | Histone deacetylases (HDACs) | Increases gene transcription |
| Methylation | Histone methyltransferases (HMTs) | Histone demethylases (KDMs) | Increases or decreases gene transcription |
| Phosphorylation | Protein kinases (PTKs) | Protein phosphatases (PPs) | Increases gene transcription & plays role in DNA repair and cell division |
| Ubiquitination | Ubiquitin ligases | Deubiquitinating enzymes (DUBs) | Increases or decreases gene transcription & plays role in DNA repair |
Acetylation

Histone acetylation, or the addition of an acetyl group to histones, is facilitated by histone acetyltransferases (HATs) which target lysine (K) residues on the N-terminal histone tail. Histone deacetylases (HDACs) facilitate the removal of such groups. The positive charge on a histone is always neutralized upon acetylation, creating euchromatin which increases transcription and expression of the target gene.[16] Lysine residues 9, 14, 18, and 23 of core histone H3 and residues 5, 8, 12, and 16 of H4 are all targeted for acetylation.[17][18]
Methylation
Histone methylation involves adding methyl groups to histones, primarily on lysine (K) or arginine (R) residues. The addition and removal of methyl groups is carried out by histone methyltransferases (HMTs) and histone demethylases (KDMs) respectively. Histone methylation is responsible for either activation or repression of genes, depending on the target site, and plays an important role in development and learning.[19] Common histone methylations include H3K9 dimethylation and H3K27 trimethylation, which are both largely associated with gene silencing, whereas H3K4, H3K36, and H3K79 methylations are associated with an increase in gene transcription.[20]
Phosphorylation
Histone phosphorylation occurs when a phosphoryl group is added to a histone. Protein kinases (PTKs) catalyze the phosphorylation of histones and protein phosphatases (PPs) catalyze the dephosphorylation of histones. Much like histone acetylation, histone phosphorylation neutralizes the positive charge on histones which induces euchromatin and increases gene expression.[21] Histone phosphorylation occurs on serine (S), threonine (T) and tyrosine (Y) amino-acid residues mainly in the N-terminal histone tails.[22]
Additionally, the phosphorylation of histones has been found to play a role in DNA repair and chromatin condensation during cell division.[23] One such example is the phosphorylation of S139 on H2AX histones, which is needed to repair double-stranded breaks in the DNA.[23]
Ubiquitination
Ubiquitination is a post-translational modification involving the addition of ubiquitin proteins onto target proteins. Histones are often ubiquitinated with one ubiquitin molecule (monoubiquitination), but can also be modified with ubiquitin chains (polyubiquitination), both of which can have variable effects on gene transcription.[24] Ubiquitin ligases add these ubiquitin proteins while deubiquitinating enzymes (DUBs) remove these groups.[25] Ubiquitination of the H2A core histone typically represses gene expression as it prevents methylation at H3K4, while H2B ubiquitination is necessary for H3K4 methylation and can lead to both gene activation or repression.[26] Additionally, histone ubiquitination is related to genomic maintenance, as ubiquitination of histone H2AX is involved in DNA damage recognition of DNA double-strand breaks.[27]
Uncommon histone modifications
Additional infrequent histone modifications and their effects are listed in the table below.[28][29][30][31][32][33][34][35][36][37][38][39][40][41][42]
| Modification | Writer(s) | Eraser(s) | Effect on DNA |
|---|---|---|---|
| O-GlcNAcylation | O-GlcNAc transferase (OGT) | O-GlcNAcase (OGA) | Increases or decreases transcription via mediation of additional histone modifications |
| Sumoylation | E3 SUMO ligases | SUMO-specific proteases | Increases or decreases transcription & plays role in DNA repair |
| ADP-ribosylation | Poly-ADP ribose polymerase 1 (PARP-1) | (Adp-ribosyl)hydrolases ARH1 & ARH3 | Decreases transcription when marking specific DNA sites for repair |
| Citrullination | Protein arginine deiminase 4 (PAD4) | No known eraser | Decreases transcription via removing methylation sites |
| Proline isomerization | Fpr4 | Fpr4 | Increases or decreases transcription via switching between H3P38 isomers (trans and cis respectively) |
O-GlcNAcylation
The presence of O-GlcNAcylation (O-GlcNAc) on serine (S) and threonine (T) histone residues is known to mediate other post-transcriptional histone modifications. The addition and removal of GlcNAc groups are performed by O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA) respectively. While our understanding of these processes is limited, GlcNAcylation of S112 on core histone H2B has been found to promote monoubiquitination of K120.[28] Similarly, OGT associates with the HCF1 complex which interacts with BAP1 to mediate deubiquitination of H2A. OGT is also involved in the trimethylation of H3K27 and creates a co-repressor complex to promote histone deacetylation upon binding to SIN3A.[29]
Sumoylation
SUMOylation refers to the addition of Small Ubiquitin-like Modifier (SUMO) proteins onto histones. SUMOylation involves covalent attachments between SUMO proteins and lysine (K) residues on histones and is carried out in three main steps by three respective enzymes: activation via SUMO E1, conjugation via SUMO E2, and ligation via SUMO E3. In humans, SUMO E1 has been identified as the heterodimer SAE1/SAE2, SUMO E2 is known as UBE2I, and the SUMO E3 role may be a multi-protein complex played by a handful of different enzymes.[30]
SUMOylation affects the chromatin status (looseness) of the histone and influences the assembly of transcription factors on genetic promoters, leading to either transcriptional repression or activation depending on the substrate.[31] SUMOylation also plays a role in the major DNA repair pathways of base excision repair, nucleotide excision repair, non-homologous end joining and homologous recombination repair. Additionally, SUMOylation facilitates error prone translesion synthesis.[32]
ADP-ribosylation
ADP-ribosylation (ADPr) defines the addition of one or more adenosine diphosphate ribose (ADP-ribose) groups to a protein.[33] ADPr is an important mechanism in gene regulation that affects chromatin organization, the binding of transcription factors, and mRNA processing through poly-ADP ribose polymerase (PARP) enzymes. There are multiple types of PARP proteins, but the subclass of DNA-dependent PARP proteins including PARP-1, PARP-2, and PARP-3 interact with the histone.[34] The PARP-1 enzyme is the most prominent of these three proteins in the context of gene regulation and interacts with all five histone proteins.[35]
Like PARPs 2 and 3, the catalytic activity of PARP-1 is activated by discontinuous DNA fragments, DNA fragments with single-stranded breaks. PARP-1 binds histones near the axis where DNA enters and exits the nucleosome and additionally interacts with numerous chromatin-associated proteins which allow for indirect association with chromatin.[34] Upon binding to chromatin, PARP-1 produces repressive histone marks that can alter the conformational state of histones and inhibit gene expression so that DNA repair can take place. Other avenues of transcription regulation by PARP-1 include acting as a transcription coregulator, regulation of RNA and modulation of DNA methylation via inhibiting the DNA methyltransferase Dnmt1.[34][36]
Citrullination

Citrullination, or deimination, is the process by which the amino acid arginine (R) is converted into citrulline. Protein arginine deiminases (PADs) replace the ketimine group of arginine with a ketone group to form the citrulline.[37] PAD4 is the deaminase involved in histone modification and converts arginine to citrulline on histones H3 and H4; because arginine methylation on these histones is important for transcriptional activation, citrullination of certain residues can cause the eventual loss of methylation, leading to decreased gene transcription;[38] specific citrullination of H3R2, H3R8, H3R17, and H3R26 residues have been identified in breast cancer cells.[39] As of research conducted in 2019, this process is thought to be irreversible.[40]
Proline Isomerization
Isomerization involves transforming a molecule so that it adopts a different structural conformation; proline isomerization plays an integral role in the modification of histone tails.[41] Fpr4 is the prolyl isomerase enzyme (PPIase) which converts the amino acid proline (P) on histones between the cis and trans conformations. While Fpr4 has catalytic activity on a number of prolines on the N-terminal region of core histone H3 (P16, P30 and P38), it most readily binds to P38.[42]
H3P38 lies near the lysine (K) residue H3K36, and changes in P38 can affect the methylation status of K36. The two possible P38 isomers available, cis and trans, cause differential effects that are opposite of each other. The cis position induces compact histones and decreases the ability of proteins to bind to the DNA, thus preventing methylation of K36 and decreasing gene transcription. Conversely, the trans position of P38 promotes a more open histone conformation, allowing for K36 methylation and leading to an increase gene transcription.[41]
Role in research
Cancer
Alterations in the functions of histone-modifying enzymes deregulate the control of chromatin-based processes, ultimately leading to oncogenic transformation and cancer.[43] Both DNA methylation and histone modifications show patterns of distribution in cancer cells.[44][45] These epigenetic alterations may occur at different stages of tumourigenesis and thus contribute to both the development and/or progression of cancer.[45]
Other Research
Vitamin B12 deficiency in mice has been shown to alter expression of histone modifying enzymes in the brain, leading to behavioral changes and epigenetic reprogramming.[46][47] Evidences also show the importance of HDACs in regulation of lipid metabolism and other metabolic pathways playing a role in the pathophysiology of metabolic disorders.[48]
See also
References
- Morgan MA, Shilatifard A (December 2020). "Reevaluating the roles of histone-modifying enzymes and their associated chromatin modifications in transcriptional regulation". Nature Genetics. 52 (12): 1271–1281. doi:10.1038/s41588-020-00736-4. PMID 33257899. S2CID 227242638.
- Vinayachandran, V; et al. (14 February 2018). "Widespread and precise reprogramming of yeast protein-genome interactions in response to heat shock". Genome Research. 28 (3): 357–366. doi:10.1101/gr.226761.117. PMC 5848614. PMID 29444801.
- McGinty RK, Tan S (March 2015). "Nucleosome structure and function". Chemical Reviews. 115 (6): 2255–2273. doi:10.1021/cr500373h. PMC 4378457. PMID 25495456.
- Kouzarides T (February 2007). "Chromatin modifications and their function". Cell. 128 (4): 693–705. doi:10.1016/j.cell.2007.02.005. PMID 17320507. S2CID 11691263.
- Sterner DE, Berger SL (June 2000). "Acetylation of histones and transcription-related factors". Microbiology and Molecular Biology Reviews. 64 (2): 435–459. doi:10.1128/MMBR.64.2.435-459.2000. PMC 98999. PMID 10839822.
- Zhang Y, Reinberg D (September 2001). "Transcription regulation by histone methylation: interplay between different covalent modifications of the core histone tails". Genes & Development. 15 (18): 2343–2360. doi:10.1101/gad.927301. PMID 11562345.
- Nowak SJ, Corces VG (April 2004). "Phosphorylation of histone H3: a balancing act between chromosome condensation and transcriptional activation". Trends in Genetics. 20 (4): 214–220. doi:10.1016/j.tig.2004.02.007. PMID 15041176.
- Shilatifard A (2006). "Chromatin modifications by methylation and ubiquitination: implications in the regulation of gene expression". Annual Review of Biochemistry. 75: 243–269. doi:10.1146/annurev.biochem.75.103004.142422. PMID 16756492.
- Sakabe K, Wang Z, Hart GW (November 2010). "Beta-N-acetylglucosamine (O-GlcNAc) is part of the histone code". Proceedings of the National Academy of Sciences of the United States of America. 107 (46): 19915–19920. Bibcode:2010PNAS..10719915S. doi:10.1073/pnas.1009023107. PMC 2993388. PMID 21045127.
- Nathan D, Ingvarsdottir K, Sterner DE, Bylebyl GR, Dokmanovic M, Dorsey JA, et al. (April 2006). "Histone sumoylation is a negative regulator in Saccharomyces cerevisiae and shows dynamic interplay with positive-acting histone modifications". Genes & Development. 20 (8): 966–976. doi:10.1101/gad.1404206. PMC 1472304. PMID 16598039.
- Hassa PO, Haenni SS, Elser M, Hottiger MO (September 2006). "Nuclear ADP-ribosylation reactions in mammalian cells: where are we today and where are we going?". Microbiology and Molecular Biology Reviews. 70 (3): 789–829. doi:10.1128/MMBR.00040-05. PMC 1594587. PMID 16959969.
- Cuthbert GL, Daujat S, Snowden AW, Erdjument-Bromage H, Hagiwara T, Yamada M, et al. (September 2004). "Histone deimination antagonizes arginine methylation". Cell. 118 (5): 545–553. doi:10.1016/j.cell.2004.08.020. PMID 15339660. S2CID 8948511.
- Wang Y, Wysocka J, Sayegh J, Lee YH, Perlin JR, Leonelli L, et al. (October 2004). "Human PAD4 regulates histone arginine methylation levels via demethylimination". Science. 306 (5694): 279–283. Bibcode:2004Sci...306..279W. doi:10.1126/science.1101400. PMID 15345777. S2CID 1579362.
- Sams, K.L; Mukai, C; Marks, B.A; Mittal, C; Demeter, E.A; Nelissen, S; Grenier, J.K; Tate, A.E; Ahmed, F; Coonrod, S.A (October 2022). "Delayed puberty, gonadotropin abnormalities and subfertility in male Padi2/Padi4 double knockout mice". Reprod Biol Endocrinol. 20 (1): 150. doi:10.1186/s12958-022-01018-w. PMC 9555066. PMID 36224627.
- Nelson CJ, Santos-Rosa H, Kouzarides T (September 2006). "Proline isomerization of histone H3 regulates lysine methylation and gene expression". Cell. 126 (5): 905–916. doi:10.1016/j.cell.2006.07.026. PMID 16959570. S2CID 17789997.
- Gräff J, Tsai LH (February 2013). "Histone acetylation: molecular mnemonics on the chromatin". Nature Reviews. Neuroscience. 14 (2): 97–111. doi:10.1038/nrn3427. PMID 23324667. S2CID 205508482.
- Roth SY, Denu JM, Allis CD (2001-06-01). "Histone acetyltransferases". Annual Review of Biochemistry. 70 (1): 81–120. doi:10.1146/annurev.biochem.70.1.81. PMID 11395403.
- Voet, D; Voet, JG (2004). Biochemistry (3rd ed.). Hoboken, N.J.: John Wiley & Sons. ISBN 978-0-471-19350-0.
- Jambhekar A, Dhall A, Shi Y (October 2019). "Roles and regulation of histone methylation in animal development". Nature Reviews. Molecular Cell Biology. 20 (10): 625–641. doi:10.1038/s41580-019-0151-1. PMC 6774358. PMID 31267065.
- "Histone Methylation - an overview | ScienceDirect Topics". www.sciencedirect.com. Retrieved 2021-12-02.
- "Histone Phosphorylation - an overview | ScienceDirect Topics". www.sciencedirect.com. Retrieved 2021-10-04.
- "Histone Phosphorylation - an overview | ScienceDirect Topics". www.sciencedirect.com. Retrieved 2021-12-02.
- Rossetto D, Avvakumov N, Côté J (October 2012). "Histone phosphorylation: a chromatin modification involved in diverse nuclear events". Epigenetics. 7 (10): 1098–1108. doi:10.4161/epi.21975. PMC 3469451. PMID 22948226.
- Guo, Hui Jun; Tadi, Prasanna (2021), "Biochemistry, Ubiquitination", StatPearls, Treasure Island (FL): StatPearls Publishing, PMID 32310512, retrieved 2021-12-02
- Smeenk G, Mailand N (2016-06-28). "Writers, Readers, and Erasers of Histone Ubiquitylation in DNA Double-Strand Break Repair". Frontiers in Genetics. 7: 122. doi:10.3389/fgene.2016.00122. PMC 4923129. PMID 27446204.
- "Histone Ubiquitination - an overview | ScienceDirect Topics". www.sciencedirect.com. Retrieved 2021-12-02.
- Ikura T, Tashiro S, Kakino A, Shima H, Jacob N, Amunugama R, et al. (October 2007). "DNA damage-dependent acetylation and ubiquitination of H2AX enhances chromatin dynamics". Molecular and Cellular Biology. 27 (20): 7028–7040. doi:10.1128/MCB.00579-07. PMC 2168918. PMID 17709392.
- Fujiki R, Hashiba W, Sekine H, Yokoyama A, Chikanishi T, Ito S, et al. (November 2011). "GlcNAcylation of histone H2B facilitates its monoubiquitination". Nature. 480 (7378): 557–560. Bibcode:2011Natur.480..557F. doi:10.1038/nature10656. PMC 7289526. PMID 22121020.
- Yang X, Qian K (July 2017). "Protein O-GlcNAcylation: emerging mechanisms and functions". Nature Reviews. Molecular Cell Biology. 18 (7): 452–465. doi:10.1038/nrm.2017.22. PMC 5667541. PMID 28488703.
- Gareau JR, Lima CD (December 2010). "The SUMO pathway: emerging mechanisms that shape specificity, conjugation and recognition". Nature Reviews. Molecular Cell Biology. 11 (12): 861–871. doi:10.1038/nrm3011. PMC 3079294. PMID 21102611.
- Lyst MJ, Stancheva I (December 2007). "A role for SUMO modification in transcriptional repression and activation". Biochemical Society Transactions. 35 (Pt 6): 1389–1392. doi:10.1042/BST0351389. PMC 2871292. PMID 18031228.
- Jalal D, Chalissery J, Hassan AH (March 2017). "Genome maintenance in Saccharomyces cerevisiae: the role of SUMO and SUMO-targeted ubiquitin ligases". Nucleic Acids Research. 45 (5): 2242–2261. doi:10.1093/nar/gkw1369. PMC 5389695. PMID 28115630.
- Ziegler M (March 2000). "New functions of a long-known molecule. Emerging roles of NAD in cellular signaling". European Journal of Biochemistry. 267 (6): 1550–1564. doi:10.1046/j.1432-1327.2000.01187.x. PMID 10712584.
- Ryu KW, Kim DS, Kraus WL (March 2015). "New facets in the regulation of gene expression by ADP-ribosylation and poly(ADP-ribose) polymerases". Chemical Reviews. 115 (6): 2453–2481. doi:10.1021/cr5004248. PMC 4378458. PMID 25575290.
- Karch KR, Langelier MF, Pascal JM, Garcia BA (November 2017). "The nucleosomal surface is the main target of histone ADP-ribosylation in response to DNA damage". Molecular BioSystems. 13 (12): 2660–2671. doi:10.1039/c7mb00498b. PMC 5702540. PMID 29058739.
- Rack JG, Ariza A, Drown BS, Henfrey C, Bartlett E, Shirai T, et al. (December 2018). "(ADP-ribosyl)hydrolases: Structural Basis for Differential Substrate Recognition and Inhibition". Cell Chemical Biology. 25 (12): 1533–1546.e12. doi:10.1016/j.chembiol.2018.11.001. PMC 6309922. PMID 30472116.
- "Citrullination - an overview | ScienceDirect Topics". www.sciencedirect.com. Retrieved 2021-12-02.
- Cuthbert GL, Daujat S, Snowden AW, Erdjument-Bromage H, Hagiwara T, Yamada M, et al. (September 2004). "Histone deimination antagonizes arginine methylation". Cell. 118 (5): 545–553. doi:10.1016/j.cell.2004.08.020. PMID 15339660. S2CID 8948511.
- Zhu D, Zhang Y, Wang S (June 2021). "Histone citrullination: a new target for tumors". Molecular Cancer. 20 (1): 90. doi:10.1186/s12943-021-01373-z. PMC 8192683. PMID 34116679.
- Darrah E, Andrade F (January 2018). "Rheumatoid arthritis and citrullination". Current Opinion in Rheumatology. 30 (1): 72–78. doi:10.1097/BOR.0000000000000452. PMC 5848217. PMID 28937414.
- Sadakierska-Chudy A, Filip M (February 2015). "A comprehensive view of the epigenetic landscape. Part II: Histone post-translational modification, nucleosome level, and chromatin regulation by ncRNAs". Neurotoxicity Research. 27 (2): 172–197. doi:10.1007/s12640-014-9508-6. PMC 4300421. PMID 25516120.
- Monneau YR, Soufari H, Nelson CJ, Mackereth CD (September 2013). "Structure and activity of the peptidyl-prolyl isomerase domain from the histone chaperone Fpr4 toward histone H3 proline isomerization". The Journal of Biological Chemistry. 288 (36): 25826–25837. doi:10.1074/jbc.M113.479964. PMC 3764789. PMID 23888048.
- Sawan, Carla; Herceg, Zdenko (2010-01-01), Herceg, Zdenko; Ushijima, Toshikazu (eds.), "3 - Histone Modifications and Cancer", Advances in Genetics, Epigenetics and Cancer, Part A, Academic Press, 70: 57–85, doi:10.1016/B978-0-12-380866-0.60003-4, ISBN 9780123808660, PMID 20920745, retrieved 2021-12-03
- Audia JE, Campbell RM (April 2016). "Histone Modifications and Cancer". Cold Spring Harbor Perspectives in Biology. 8 (4): a019521. doi:10.1101/cshperspect.a019521. PMC 4817802. PMID 27037415.
- Kurdistani SK (July 2007). "Histone modifications as markers of cancer prognosis: a cellular view". British Journal of Cancer. 97 (1): 1–5. doi:10.1038/sj.bjc.6603844. PMC 2359665. PMID 17592497.
- Ghosh S, Sinha JK, Khandelwal N, Chakravarty S, Kumar A, Raghunath M (September 2020). "Increased stress and altered expression of histone modifying enzymes in brain are associated with aberrant behaviour in vitamin B12 deficient female mice". Nutritional Neuroscience. 23 (9): 714–723. doi:10.1080/1028415X.2018.1548676. PMID 30474509. S2CID 53785219.
- Saraswathy KN, Ansari SN, Kaur G, Joshi PC, Chandel S (April 2019). "Association of vitamin B12 mediated hyperhomocysteinemia with depression and anxiety disorder: A cross-sectional study among Bhil indigenous population of India". Clinical Nutrition ESPEN. 30: 199–203. doi:10.1016/j.clnesp.2019.01.009. PMID 30904222. S2CID 85497723.
- Ferrari A, Fiorino E, Giudici M, Gilardi F, Galmozzi A, Mitro N, et al. (November 2012). "Linking epigenetics to lipid metabolism: focus on histone deacetylases". Molecular Membrane Biology. 29 (7): 257–266. doi:10.3109/09687688.2012.729094. PMID 23095054. S2CID 39956339.