Immune complex
An immune complex, sometimes called an antigen-antibody complex or antigen-bound antibody, is a molecule formed from the binding of multiple antigens to antibodies.[1] The bound antigen and antibody act as a unitary object, effectively an antigen of its own with a specific epitope. After an antigen-antibody reaction, the immune complexes can be subject to any of a number of responses, including complement deposition, opsonization,[2] phagocytosis, or processing by proteases. Red blood cells carrying CR1-receptors on their surface may bind C3b-coated immune complexes and transport them to phagocytes, mostly in liver and spleen, and return to the general circulation.
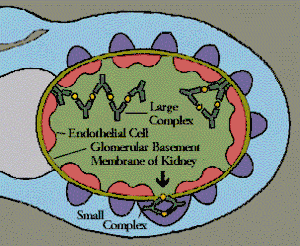
The ratio of antigen to antibody determines size and shape of immune complex.[3] This, in turn, determines the effect of the immune complex. Many innate immune cells have FcRs, which are membrane-bound receptors that bind the constant regions of antibodies. Most FcRs on innate immune cells have low affinity for a singular antibody, and instead need to bind to an immune complex containing multiple antibodies in order to begin their intracellular signaling pathway and pass along a message from outside to inside of the cell.[3] Additionally, the grouping and binding together of multiple immune complexes allows for an increase in the avidity, or strength of binding, of the FcRs. This allows innate immune cells to get multiple inputs at once and prevents them from being activated early.[3]
Immune complexes may themselves cause illness when they are deposited in organs, for example, in certain forms of vasculitis. This is the third form of hypersensitivity in the Gell-Coombs classification, called type III hypersensitivity.[4] Such hypersensitivity progressing to disease states produces the immune complex diseases.
Immune complex deposition is a prominent feature of several autoimmune diseases, including rheumatoid arthritis, scleroderma and Sjögren's syndrome.[5][6] An inability to degrade immune complexes in the lysosome and subsequent accumulation on the surface of immune cells has been associated with systemic lupus erythematosus.[7][8]
Functions
Regulation of antibody production
Immune complexes can also play a role in the regulation of antibody production. B cells express B-cell receptors (BCRs) on their surfaces and antigen binding to these receptors begins a signaling cascade that leads to activation. B cells also express FcγRIIb, low affinity receptors specific to the constant region of IgG, on their surfaces. IgG immune complexes are the ligand for these receptors and immune complex binding to these receptors induces apoptosis, or cell death. After B cells are activated, they differentiate into plasma cells and cease to express BCR but continue to express FcγRIIb, which allows IgG immune complexes to regulate IgG production via negative feedback and prevent uncontrolled IgG production.[9]
Activation of dendritic cells and macrophages
Immune complexes, particularly those made of IgG, also play a variety of roles in the activation and regulation of phagocytes, which include dendritic cells (DCs) and macrophages. Immune complexes are better at inducing DC maturation than an antigen on its own.[10] Again, the low affinity of many FcγR for IgG means that only immune complexes, not single antibodies, can induce the FcγR’s signaling cascade. When compared to single antibodies binding to FcγRs, immune complexes binding to FcγRs cause significant changes in internalization and processing of antigen, maturation of the vesicles containing the internalized antigen, and activation in DCs and macrophages.[11] There are multiple classes of macrophages and DCs that express different FcγRs, which have differing affinities for single antibodies and immune complexes.[11] This allows the response of the DC or macrophage to be tuned precisely, subsequently tuning the level of IgG. These diverse FcγRs cause different responses in their DCs or macrophages by initiating different signaling pathways that can either activate or inhibit cellular functions.[11] The binding of the immune complex to the DC’s membrane-bound receptor and internalization of the immune complex and receptor begins the process of antigen presentation, which allows the DC to activate T cells. Via this process, immune complexes cause enhanced T cell activation.[11]
Elimination of opsonized immune complexes
Type I FcγRs, another type of IgG constant region receptor, can bind to IgG immune complexes and lead to the elimination of the opsonized complex. Immune complexes bind to multiple type I FcγRs, which cluster on the cell surface and begin the ITAM signaling pathway. This signaling pathway involves the phosphorylation of specific amino acids within a sequence of proteins and eventually leads to elimination of opsonized immune complex.[9]
References
- Cush, John; Kavanaugh, Arthur; Stein, Charles (2005). Rheumatology: Diagnosis and Therapeutics. Lippincott Williams & Wilkins. p. 78. ISBN 9780781757324.
- Goldsby, Richard (2002). Immunology. Macmillan. p. 381. ISBN 9780716749479.
- Lu, Lenette L.; Suscovich, Todd J.; Fortune, Sarah M.; Alter, Galit (January 2018). "Beyond binding: antibody effector functions in infectious diseases". Nature Reviews Immunology. 18 (1): 46–61. doi:10.1038/nri.2017.106. ISSN 1474-1733. PMC 6369690. PMID 29063907.
- Barret, James (1980). Basic Immunology and its Medical Application (2 ed.). St.Louis: The C.V. Mosby Company. ISBN 0-8016-0495-8.
- Lawley, Thomas; Moustopoulos, Haralampos (1979). "Demonstration of Circulating Immune Complexes in Sjögren's Syndrome". Journal of Immunology. The American Association of Immunologists. 123 (3): 1382–7. doi:10.4049/jimmunol.123.3.1382. PMID 469255. S2CID 31511399.
- Wallace, Daniel, ed. (2004). The New Sjogren's Syndrome Handbook. Oxford University Press. p. 68. ISBN 9780198038481.
- Monteith, Andrew J.; Kang, SunAh; Scott, Eric; Hillman, Kai; Rajfur, Zenon; Jacobson, Ken; Costello, M. Joseph; Vilen, Barbara J. (2016-04-12). "Defects in lysosomal maturation facilitate the activation of innate sensors in systemic lupus erythematosus". Proceedings of the National Academy of Sciences. 113 (15): E2142–E2151. Bibcode:2016PNAS..113E2142M. doi:10.1073/pnas.1513943113. ISSN 0027-8424. PMC 4839468. PMID 27035940.
- Kang, SunAh; Rogers, Jennifer L.; Monteith, Andrew J.; Jiang, Chuancang; Schmitz, John; Clarke, Stephen H.; Tarrant, Teresa K.; Truong, Young K.; Diaz, Marilyn; Fedoriw, Yuri; Vilen, Barbara J. (2016-05-15). "Apoptotic Debris Accumulates on Hematopoietic Cells and Promotes Disease in Murine and Human Systemic Lupus Erythematosus". The Journal of Immunology. 196 (10): 4030–4039. doi:10.4049/jimmunol.1500418. ISSN 0022-1767. PMC 4868781. PMID 27059595.
- Bournazos, Stylianos; Wang, Taia T.; Dahan, Rony; Maamary, Jad; Ravetch, Jeffrey V. (2017-04-26). "Signaling by Antibodies: Recent Progress". Annual Review of Immunology. 35 (1): 285–311. doi:10.1146/annurev-immunol-051116-052433. ISSN 0732-0582. PMC 5613280. PMID 28446061.
- Nelson, Nicole L.J.; Zajd, Cheryl M.; Lennartz, Michelle R.; Gosselin, Edmund J. (November 2019). "Fcγ receptors and toll-like receptor 9 synergize to drive immune complex-induced dendritic cell maturation". Cellular Immunology. 345: 103962. doi:10.1016/j.cellimm.2019.103962. PMC 6892604. PMID 31582169.
- Guilliams, Martin; Bruhns, Pierre; Saeys, Yvan; Hammad, Hamida; Lambrecht, Bart N. (February 2014). "The function of Fcγ receptors in dendritic cells and macrophages". Nature Reviews Immunology. 14 (2): 94–108. doi:10.1038/nri3582. ISSN 1474-1733. PMID 24445665. S2CID 11733324.