Labyrinthula
The genus Labyrinthula is part of the protist group Labyrinthulomycetes and contains thirteen species. The major feature of this genus is the formation of an ectoplasmic net secreted by specialized organelles called bothrosomes which surrounds the colony, which is also used by Labyrinthula for moving. The protist reproduces by zoosporulation as it sets some flagellated spores free from a sporangium. One of the flagella of the zoospores has stiff tripartite hairs (mastigonemes) - the defining characteristic of the stramenopiles.
| Labyrinthula | |
|---|---|
| Scientific classification | |
| Domain: | |
| (unranked): | |
| Superphylum: | |
| Phylum: | |
| Class: | Labyrinthulae |
| Order: | |
| Family: | Labyrinthulidae |
| Genus: | Labyrinthula |
The interest in Labyrinthula arose as it has been identified as the cause of the "wasting disease", which led to the death of more than 90% of the seagrass population of the North Atlantic coast in the early 1930s.[1]
Etymology
A labyrinth is a synonym of a maze, reflecting the ectoplasmic net, which gives the protist a net-like shape which resembles a maze. The word comes from Ancient Greek λαβύρινθος (labúrinthos, “maze”).
History of Knowledge
Cienkowski was the first person who published on Labyrinthula in 1867.[2] He studied primarily the morphology, and he identified two species: Labyrinthula vitelli and Labyrinthula macrocystis.
In 1967, Pokorny published the first review over Labyrinthula and counted ten marine species, two freshwater species and one terrestrial species.[3]
Before being considered as protists and Stramenopiles,[4] Labyrinthula had been classified in the Mycetozoa,[5] the slime molds[6] and the Rhizopodia.[7]
Interest in Labyrinthula started when it was defined as the cause of the "wasting disease". An epidemic happened in the 1930s in the North American and European coasts, killing over 90% of the seagrass (Zostera marina) population.[1] The first description of similar symptoms in terrestrial grasses happened in 1995 in California. The target organism is turfgrass and the pathogen identified by the Koch's postulates is L. terrestris.[8] Both seagrass and turfgrass can be infected by organisms of the Labyrinthula genera, but from different species, one marine and one terrestrial one, respectively.
Habitat and Ecology
The habitat of Labyrinthula is very diverse, they are found in marine and freshwater environments as well as in terrestrial environments all over the world. Labyrinthula have been found to live as pathogens, commensals or mutualists on several host organisms. One example is the endosymbiont Labyrinthula sp. inside Thecamoeba hilla, which seem to live as mutualists symbionts.[9]
Description of the Organism
Morphology and Anatomy
The morphology of a single cell of Labyrinthula is not unique and varies a lot between the different species. The cells can be spindle-shaped like L. macrocystis, spherical or ovoid to name only a few examples. The spindle shape is due to the microtubules of the cytoskeleton.[10] There is no mean size of one cell, because of the diversity between the species. An approximation can still be made at 8-30 γm in length and 1.5-8 γm in width.[3] The general morphology of the cell contain one central nucleus, the smooth endoplasmatic reticulum, mitochondria with tubular cristae, numerous lipid-composed granules and two large Golgi complexes.[9] Labyrinthula are non-photosynthetic and are usually translucent white, but some yellow strains have been observed.[11]
Ectoplasmic Net
The defining characteristic of the genera Labyrinthula is the formation of an ectoplasmic net around the cells and embedding the whole colony. The ectoplasmic net is secreted and attached to the cell by specialized organelles called segenetosome or bothrosomes. A bothrosome is an electron-opaque organelle, which prevents the leaking of the organelles into the net. The etymology of "bothrosome" and "sagenetosome" originated from bothros: hole and soma: body, as well as from sagena: net, genetes: ancestor and soma: body respectively. The net is composed of secreted ectoplasm and is delimited by a plasma membrane. It lacks a cell wall and contains no organelles. By forming long filaments, the ectoplasmic net allows the colony to attach to surfaces and it secretes digestive enzymes for absorptive nutrition. These enzymes can be surface-bound or secreted into the medium to help the digestion of organic substances. Individual cells use the ectoplasmic net for movement by gliding inside it. They move in all directions, but they tend to go towards the periphery of the ectoplasmic net to enlarge the net and thus increase its surface area.[12][13]
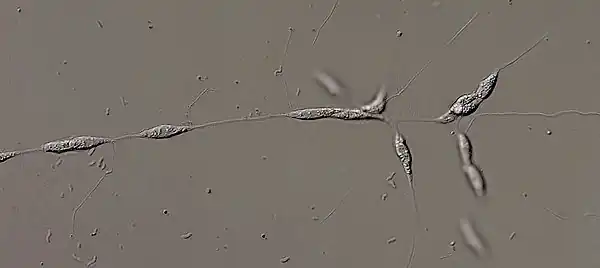
Cell Wall
Despite the lack of cell wall of the ectoplasmic net, each individual cell is surrounded by a cell wall located close to the cell membrane and composed of a single layer of Golgi-derived circular scales, which overlap over a few nanometers, but do not fuse. The main components of the cell wall are fucose or galactose-derived substances.[12]
Life Cycle
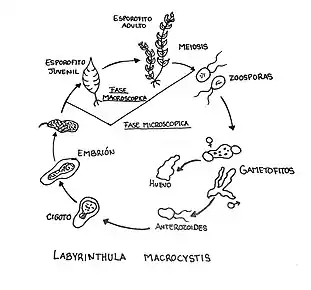
The reproduction of Labyrinthula occurs by zoosporulation, no sexual reproductive cells or structures have been identified yet. First, vegetative cells aggregate inside the ectoplasmic net and form yellow to orange networks in different areas of the net. The cells in these aggregates are compressed together and the boundaries blur, which leads to a plasmodium-like appearance. The vegetative cells then round up and enlarge to form a presporangium. A presporangium is delimited by an envelope of scales, which build a rather thick membrane. The presporangium is further divided into sporangia, and in some species into a sori,[14] until each of them contain eight zoospores. The zoospores are released directly into the ectoplasmic net, where they swim away from the sporangium.[15]
The morphology of the zoospore is similar to a "standard" stramenopile cell. Zoospores contain an eyespot and two flagella. Only the anterior flagellum is covered with mastigonemes and the posterior flagellum propels the cell through the ectoplasmic net. Both flagella are inserted laterally and medially. After about 24 hours, the zoospores lose their flagella and round up to finally differentiate into vegetative spindle cells.[15][14] Vegetative cells stop gliding through the net in older cultures or under bad conditions. Only few of the older cells autolyze, but most of them round up.[16]
Vegetative cells multiply mostly by mitosis. Some features of their binary fission are the de novo synthesis of the bothrosome and the cytokinesis, that occurs by vesicle accumulation and fusion.[10]
Practical Importance
Labyrinthula have awoken the interest of scientists by being the cause of the "wasting disease" of the seagrass on the North American and European coasts in the 1930s. Since then, several pathogenic species have been identified which mostly live in marine water.
Studies testing the virulence of the protist in seagrasses showed a low virulence of Labyrinthula. The protist often lives inside the multicellular organism, but does not initiate any pathogenic event. Pathogenesis may be triggered by biotic or abiotic stress of the higher organism, like a large increase in temperature or a decrease in light exposure for example.[17]
The most famous parasitic symbiont is L. zosterae, which colonizes marine seagrasses referring to the event mentioned above. Typical features of the "wasting disease" are a discolouration of the leaves, due to the destruction of the chloroplasts in epidermis, mesophyll and lesion formation. The second stage is the occurrence of brown and black blotches.[13] The infection and the following consequences are a concern, because of the ecological and economical importance of the seagrass in the marine environment.[18] The infection is transferred by direct contact between the target organisms and correlates with the salinity of the water.[18]
Another parasitic Labyrinthula is L. terrestris, which is a terrestrial parasite of turf grasses. It has been identified by the Koch's postulates as responsible for the "rapid blight" killing infected turf grasses.[8] Some other symbiont hosts of Labyrinthula are Chaetomorpha media, a green algae[19] and Thecamoeba hilla where they live as mutualistic symbiont.[9]
List of Species
| Species | Discover | Year |
|---|---|---|
| L. macrocystis [2] | CIenkowski | 1867 |
| L. vitellina [2] | Cienkowski | 1867 |
| L. cienkowskii [5] | Zopf | 1892 |
| L. zopfii | Dangeard | 1910 |
| L. chattonii [20] | Dangeard | 1932 |
| L. valkanovii [21] | Karling | 1944 |
| L. algeriensis [22] | Hollande & Enjumet | 1955 |
| L. roscoffensis [23] | Chadefaud | 1956 |
| L. spp / L. zosterae | Watson | 1957 |
| L.sp Type LX | Watson | 1957 |
| L. sp Type SELX | Watson | 1957 |
| L. coenocystis [24] | Schmoller | 1960 |
| L. terrestris [8] | Bigelow | 2005 |
References
- Muehlstein, Lisa K.; Porter, David; Short, Frederick T. (1991-01-01). "Labyrinthula zosterae sp. nov., the Causative Agent of Wasting Disease of Eelgrass, Zostera marina". Mycologia. 83 (2): 180–191. doi:10.2307/3759933. JSTOR 3759933.
- Cienkowski, L. (1867-12-01). "Ueber den Bau und die Entwickelung der Labyrinthuleen". Archiv für Mikroskopische Anatomie (in German). 3 (1): 274–310. doi:10.1007/BF02960460. ISSN 0176-7364. S2CID 83753548.
- Pokorny, Kathryn Stein (1967-11-01). "Labyrinthula". The Journal of Protozoology. 14 (4): 697–708. doi:10.1111/j.1550-7408.1967.tb02065.x. ISSN 1550-7408.
- Patterson, D. J. (1989). "Stramenopiles, chromophytes from a protistan perspectives". The Chromaphyte Algae Problems and Perspectives: 357–379.
- Zopf, W. (1892). "Zur Kenntnis der Labyrinthuleen einer Familie der Myceozoen". Beitr. Hysiol. Morphol. Nied. Organ. 2: 36–48.
- Tyler., Bonner, John (2015-12-08). Cellular Slime Molds. Princeton University Press. ISBN 9781400876884. OCLC 957504952.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Honigberg, B. M.; Balamuth, W.; Bovee, E. C.; Corliss, J. O.; Gojdics, M.; Hall, R. P.; Kudo, R. R.; Levine, N. D.; Lobblich, A. R. (1964-02-01). "A Revised Classification of the Phylum Protozoa*". The Journal of Protozoology. 11 (1): 7–20. doi:10.1111/j.1550-7408.1964.tb01715.x. ISSN 1550-7408. PMID 14119564.
- Bigelow, D. M.; Olsen, M. W.; Gilbertson, R. L. (2005-03-01). "Labyrinthula terrestris sp. nov., a new pathogen of turf grass". Mycologia. 97 (1): 185–190. doi:10.1080/15572536.2006.11832852. ISSN 0027-5514. PMID 16389970. S2CID 28774010.
- Dyková, Iva; Fiala, Ivan; Dvořáková, Helena; Pecková, Hana (2008-11-01). "Living together: The marine amoeba Thecamoeba hilla Schaeffer, 1926 and its endosymbiont Labyrinthula sp". European Journal of Protistology. 44 (4): 308–316. doi:10.1016/j.ejop.2008.04.001. PMID 18619822.
- Porter, David (1972-12-01). "Cell division in the marine slime mold, Labyrinthula sp., and the role of the bothrosome in extracellular membrane production". Protoplasma. 74 (4): 427–448. doi:10.1007/BF01281960. ISSN 0033-183X. S2CID 41333647.
- Sullivan, Brooke K.; Robinson, Katie L.; Trevathan-Tackett, Stacey M.; Lilje, Erna S.; Gleason, Frank H.; Lilje, Osu (2017-02-01). "The First Isolation and Characterisation of the Protist Labyrinthula sp. in Southeastern Australia". Journal of Eukaryotic Microbiology. 64 (4): 504–513. doi:10.1111/jeu.12387. hdl:11343/292461. ISSN 1550-7408. PMID 28004878. S2CID 162410.
- Thomas., Moss, Stephen (1986-01-01). The biology of marine fungi. Cambridge University Press. ISBN 9780521308991. OCLC 610636209.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Raghukumar, Seshagiri (2002). "Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids)". European Journal of Protistology. 38 (2): 127–145. doi:10.1078/0932-4739-00832.
- W., Dick, Michael (2010-01-01). Straminipilous Fungi. Springer Netherlands. ISBN 9789401597333. OCLC 961064561.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Perkins, Frank O.; Amon, James P. (1969-05-01). "Zoosporulation in Labyrinthula sp.; an Electron Microscope Study1". The Journal of Protozoology. 16 (2): 235–257. doi:10.1111/j.1550-7408.1969.tb02265.x. ISSN 1550-7408.
- Babtsch, G.; Bartsch, G. (1971-01-01). "Cytologische Beobachtungen an Labyrinthula coenocystis SCHMOLLER bei verschiedenen Kulturbedingungen". Zeitschrift für Allgemeine Mikrobiologie. 11 (2): 79–90. doi:10.1002/jobm.19710110202. ISSN 1521-4028.
- Brakel, Janina; Werner, Franziska Julie; Tams, Verena; Reusch, Thorsten B. H.; Bockelmann, Anna-Christina (2014-04-01). "Current European Labyrinthula zosterae Are Not Virulent and Modulate Seagrass (Zostera marina) Defense Gene Expression". PLOS ONE. 9 (4): e92448. Bibcode:2014PLoSO...992448B. doi:10.1371/journal.pone.0092448. ISSN 1932-6203. PMC 3972160. PMID 24691450.
- Muehlstein, LK (1989). "Perspectives on the wasting disease of eelgrass Zostera marina". Diseases of Aquatic Organisms. 7: 211–221. doi:10.3354/dao007211.
- Raghukumar, C (1987). "Fungal parasites of marine algae from Mandapam (South India)". Diseases of Aquatic Organisms. 3: 137–145. doi:10.3354/dao003137.
- Dangeard, P.A. (1932). "Observations sur la famille des Labyrin- thulees et sur quelques parasites des Cladophora". Le Botaniste. 24: 217.
- Karling, John S. (1944-01-01). "Phagomyxa algarum N. Gen., N. Sp., An Unusual Parasite with Plasmodiophoralean and Proteomyxean Characteristics". American Journal of Botany. 31 (1): 38–52. doi:10.2307/2437666. JSTOR 2437666.
- Hollande, A.; Enjumet, M. (1955). "Sur l'évolution et la systématique des Labyrinthulidae; étude de Labyrinthula algeriensis nov. sp". Annls. Sci. Nat. Zool. Biol. Anim. 17: 357–368.
- Chadefaud, M. (1956). "Sur un Labyrinthula de Roscoff". C. R. Acad. Sci. 243: 1794–1797.
- Schmoller, H. (1960). "Kultur und Entwicklung yon Labyrinthula coenocystis n. sp". Arch. Mikrobiol. 36 (4): 365–372. doi:10.1007/bf00418643. S2CID 6841006.