Larinioides sclopetarius
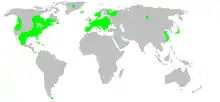
Larinioides sclopetarius, commonly called bridge-spider [1]or gray cross-spider, is a relatively large orb-weaver spider with Holarctic distribution. These spiders originated in Europe, have been observed as south as the Mediterranean Coast and as north as Finland, and have been introduced to North America. They are often found on bridges, especially near light and over water.[2] The species tends to live on steel objects and is seldom seen on vegetation.[3][4] Females reach a body length of 10–14mm, and males 8–9mm.[2] Their orb webs can have diameters of up to 70 cm.
| Larinioides sclopetarius | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Subphylum: | Chelicerata |
| Class: | Arachnida |
| Order: | Araneae |
| Infraorder: | Araneomorphae |
| Family: | Araneidae |
| Genus: | Larinioides |
| Species: | L. sclopetarius |
| Binomial name | |
| Larinioides sclopetarius | |
 | |
| Synonyms | |
|
Araneus sclopetarius | |
L. sclopetarius is attracted to light. Spiders found near light sources may be in better condition and have greater reproductive success than spiders living in unlit areas.[5] Most of these lighted areas are found in cities or other metropolitan areas. As a result, many urban areas have become saturated with these spiders. As many as 100 of these spiders can be found in a square meter in optimal feeding locations.[6]
L. sclopetarius often hide during the day, and wait for prey in the center of their web at night. In high-density populations, spiders may remain sedentary to protect their territories. Mature females and juvenile spiders will often build webs. However, adult males generally inhabit adult female webs. Males can be found mostly during summer, while females are active until November in Central Europe.
Taxonomy
The common name “bridge-spider” is attributed to its preferential habitat of building webs on bridges.[7]
Lariniodes sclopetarius was first recorded by Carl Clerck in 1757.[8] The genus Lariniodes indicates that they create large noticeable orb webs.
There has been some dispute in how the spider should be classified. The spider has been proposed as a synonym for L. cornutus due to their similar male pedipalps. However, their habitats vary greatly. This finding has been used as evidence that these two species are distinct.[8]
Description
L. sclopetarius exhibit a slight sexual dimorphism, where females are heavier than males. Females typically weigh around 60 mg, whereas males weigh around 38 mg.[9] However, males may be slightly larger than females. Females can range from 4.5- 6.25 mm in length. Males vary from 4.25 mm -7 mm in length.[9]
L. sclopetarius differs from its close relatives, L. patagiatus and L. cornutus, with a few defining characteristics. Unlike the latter two species, L. sclopetarius has white hairs that provide a silhouette for their heads and dark markings on its abdomen.[10]
Plasticity
L. sclopetarius shows high plasticity levels based on resource availability.[11] These spiders exhibit variability in growth and weight gain between molting-periods. Their intermolt periods have a broader range of values than other spiders that inhabit urban environments such as Zygiella x‐notata.
Additionally, L. sclopetarius can alter its growth rate without increasing its risk of mortality.[11] In resource-abundant areas, spiders have exhibited accelerated maturation. Their fast growth rates may allow them to colonize areas that can accommodate them.
Habitat and Distribution
In North America L. sclopetarius is most common in the Great Lakes states, but can be found throughout the country. It is mostly found near buildings.[7] L. sclopetarius is also found in Central Europe, often near water.[12] They are also most commonly found near artificial sources of light near water bodies such as bridges and boats. They often aggregate in high densities near light sources. The lights tend to attract more insects which increases the spider's prey capture.[13] Their light-seeking behaviors may have some genetic basis.
In these feeding-locations, adult females tend to occupy the best foraging areas. Juveniles and immature spiders are relegated to worse feeding areas when competition is high.[14] These behaviors may be observed because to juveniles are unable to compete for better territories. However, when these spiders mature, they often search for more illuminated areas to create their webs and lay their eggs.
Territoriality
L. sclopetarius is not a social spider.[15] However, they often build webs next to each other.[6] Females exhibit territorial defense of their webs from intruders, including other members of the same species. In high population densities, females exhibit more aggressive behavior to conspecifics due to the shortages of territories.[15]
Diet
These spiders are primarily nocturnal foragers. Their prey capture fluctuates with the seasons. Their prey capture is highest in the summer months, declining in the spring and fall. Chironomids consist of a significant portion of their diet.[13] These small flies may comprise up to 94% of the spiders’ diets.[16] Their prey's average size ranges from 1.2 to 6.8 mm.
Webs
L. sclopetarius creates circular orb webs unlike other orb-web spiders that construct elliptical orb webs.[17] Additionally, their orb-webs change in shape as the spider ages. As the spider matures, the adhesive web's lower-section will continue to increase whereas the web's upper section will become proportionally smaller.[18] This discrepancy in web-size becomes more prominent as the spider gets larger. Spiders in high-prey areas create webs with larger capture areas.[18] These trends differ from other spiders such as A. keyserlingi where satiated spiders created smaller webs.
L. sclopetarius creates webs near sources of light. The part of the web adjacent to the light captures more prey than parts of the web farther away from the light .[19]
When resting in webs, these spiders tend to orient their posteriors in the direction of the wind. When the direction of the wind changes, the spider attempts to position itself back towards it. This behavior may have evolved to reduce the spider's risk of being blown off the web's hub. Adhering to webs may be a challenge for L. sclopetarius in urban environments due to the lack of areas that can accommodate stable web sites under windy conditions.[19]
Web-Type
Adults create asymmetrical webs. The outer frame threads are generally similar in structure. The hubs, which are centers of the web, are also comparable to each other. The difference lies in the radii of the capture threads.[18] The lower region radii of the capture threads are larger than the upper regions of the web. The web's unequal sizes may be due to the spider's easier accessibility to the captured prey in the lower regions of the web.[18] This capture web asymmetry increases with spider weight and becomes more pronounced as the spiders age. Unlike adults, juveniles create symmetrical webs.
Web-Construction
The mean mesh height - the distance between different capture threads - is approximately 2 mm.[16] Unlike many other spiders, L. sclopetarius does not follow the trend of greater mesh height equating to capturing larger prey. One explanation may be the lack of prey diversity. Regardless of mesh height, L. sclopetarius primarily targets smaller dipterans.[20]
_female%252C_Arnhem%252C_the_Netherlands_-_2.jpg.webp)
Reproduction and Life-cycle
L. sclopetarius spiders live for approximately 1.5 years under optimal conditions. Females have an above average reproductive ability producing up to 15 egg sacs.[11] Males typically grow faster than females and mature into adults up to a month earlier.[9]
Unlike many spiders, development is independent of the seasons. Adult spiders may mature at any point during the year, however the greatest concentration of mature spiders occurs in late Summer.[11]
Mating
Assortative mating may be at play, as aggressive males and females are more likely to mate with each other. Additionally, non-aggressive individuals are more likely to mate with each other.[22] Aggression may be selected for as it may be important for securing the best sites for web building, which are around light sources. In some instances, groups of aggressive spiders may have lower mortality rates than mixed groups with non-aggressive and aggressive spiders.[22]
Aggressive males tend to create more fit males. Female reproductive success was partially dependent on female size rather than aggressiveness.[22]
Sexual Cannibalism
Females may eat males when resources are scarce. Females that consume insects generally absorb more lipids, which provide a more efficient energy-source. Sexual cannibalism does not seem to affect spider egg sac mass and clutch size because protein and lipid diets had elicited similar results.[12] However, when food sources are scarce, spiderlings from females consuming high lipid diets may have better survival rates than spiderlings from females consuming high protein diets. Males may be seen as last-resort prey where there are no better options.
Spiderlings from mothers who consumed male L. sclopetarius were observed spinning their webs sooner than mothers that solely ate insects.[12] This observed behavior may either be due to the male providing necessary proteins for silk production or an environmental stressor that encouraged web building to compete in resource-scarce settings.
Sexual Dimorphism
This spider follows Rensch's rule which states that male size is more variable than female size. There is only a small sexual dimorphism between males and females. Females grow slower and have more instars than males. As a result, they become heavier than males. Males grow faster and weigh approximately 40% of females. They also have longer legs and are larger than female spiders.[9] These longer leg lengths may be due to mating advantages due to female choice or male competition.
Diet may be related to male spider size. Poorer diets are associated with male spiders that possess shorter legs. Female spider size was unaffected by the quality of the diet.[9] Female size may be undergoing stabilizing selection, which can explain the lack of variance in size despite diet changes.
Social Behavior
Female spiders live independently, defending their own webs. However, male spiders may have a kleptoparasitic relationship with a female. They often choose to live on a female's web and steal their prey.[15]
L. sclopetarius exhibits high levels of activities in unfamiliar environments, which may have contributed to their widespread colonization of urban areas.[6] In experimental conditions, they tend to move and explore new settings more than their other urban counterparts Zygiella x-notata.
L. sclopetarius exhibits many aggressive behaviors such as chasing and attacking conspecifics. If individuals are in the same web, they may engage in web-shaking contests. These aggressive behaviors may be genetically inherited. Males are generally more aggressive than females (https://link.springer.com/article/10.1007/s00265-017-2353-x).[6]
L. sclopetarius typically positions itself on the lower areas of the web. This can be seen as a defensive behavior as it allows the spider to easily escape from the web from predators by using a safety line.[19]
Physiology
Locomotion
L. sclopetarius moves between areas using a ballooning technique where the spider releases threads into the wind to travel. This method allows the spider to travel to preferable feeding areas.[14]
Glands and Toxins
L. sclopetarius secretes anti-adhesive compounds that prevent its legs from sticking to their capture-threads.[23] The mechanism for how these spiders develop and secrete this protection is currently unknown.
Enemies
Phalacrotophora epeirae is one predator that consumes L. sclopetarius eggs.[24]
Trypoxylon attenuatum is a species of spider hunting wasp exhibited to hunt L. sclopetarius.[25] These wasps are normally found in Southern Europe and prey on both foraging and sedentary spiders. T. attenuatum will paralyze spiders and bring them back to their nests. Once at the nest, L. sclopetarius may have an egg laid in it and serve as a food source for the wasp's larvae.
Interactions with Humans and Animals
L. sclopetarius has been found in urban environments aggregating around light sources.[15] In Finland, they have been found in boats and boathouses.[10] These spiders can migrate via boats which have caused them to be found in isolated islands such as the Åland Islands and spread over much of coastal Europe. They pose a hazard to local businesses as restaurant boats may experience a decline in productivity due to their web's frequent presence driving away customers.
The annual migration of Larinioides sclopetarius is notable in Chicago, where ballooning spiders attach themselves to high rise buildings.[26][27] L. sclopetarius webs serve as a balloon to move through air currents till they find an ideal location to construct their new webs. This usually causes panic within people. However, this is a common occurrence especially in high rise buildings because these spiders prefer higher ground. Because of this, L. sclopetarius prefers to reside along hotel windows, due to the added benefit of bright lights for insect prey to get trapped in their webs.
Bites to Humans
Even when these spiders are present together in large numbers, they typically do not bite humans. L. sclopetarius is venomous, however the effect of the venom ranges from the potency of a mosquito-bite to the sting of a honeybee. If they do bite, it is when their webs are threatened, and the bite is usually superficial and heals quickly without much need for medical help.[26]
References
- šEstáKová, Anna; Marusik, Yuri M.; Omelko, Mikhail M. (2014-11-11). "A revision of the Holarctic genus Larinioides Caporiacco, 1934 (Araneae: Araneidae)". Zootaxa. 3894 (1): 61. doi:10.11646/zootaxa.3894.1.6. ISSN 1175-5334.
- Roberts, Michael J. (1996) Collins Field Guide: Spiders of Britain and Northern Europe, AND NORTHERN MICHIGAN Collins, ISBN 978-0-00-219981-0, pp. 321–2
- Nieuwenhuys, Ed (17 November 2013). "Orb web spiders or orb-weavers". Retrieved 3 September 2014.
- šEstáKová, Anna; Marusik, Yuri M.; Omelko, Mikhail M. (2014-11-11). "A revision of the Holarctic genus Larinioides Caporiacco, 1934 (Araneae: Araneidae)". Zootaxa. 3894 (1): 61. doi:10.11646/zootaxa.3894.1.6. ISSN 1175-5334.
- Gomes, Dylan G.E. (2020-03-17). "Orb-weaving spiders are fewer but larger and catch more prey in lit bridge panels from a natural artificial light experiment". PeerJ. 8: e8808. doi:10.7717/peerj.8808. ISSN 2167-8359. PMC 7083158. PMID 32211243.
- Kralj-Fišer, Simona; Hebets, Eileen A.; Kuntner, Matjaž (2017-07-25). "Different patterns of behavioral variation across and within species of spiders with differing degrees of urbanization". Behavioral Ecology and Sociobiology. 71 (8): 125. doi:10.1007/s00265-017-2353-x. ISSN 1432-0762. S2CID 23043867.
- Levi, Herbert (1974). "The orb-weaver genera Araniella and Nuctenea (Araneae: Araneidae)". Bulletin of the Museum of Comparative Zoology at Harvard College. 146: 291–316 – via BHL.
- Breitling, Rainer; Bauer, Tobias (2015-11-01). "Remarks on Synonyms of European Larinioides Species (Arachnida: Araneae: Araneidae)". Arachnology. 16 (9): 305–310. doi:10.13156/arac.2015.16.9.305. ISSN 2050-9928. S2CID 85900771.
- Kleinteich, Anja (2014). "Evidence for Rensch's rule in an orb-web spider with moderate sexual size dimorphism". Evolutionary Ecology Research. 12: 667–683.
- Fritzen, Niclas (2005). "Larinioides sclopetarius and Agalenatea redii (Araneae: Araneidae) - Two spider species new to Finland". Memoranda - Societatis Pro Fauna et Flora Fennica. 81: 108–110 – via ResearchGate.
- Kleinteich, Anja; Schneider, Jutta M. (February 2011). "Developmental strategies in an invasive spider: constraints and plasticity". Ecological Entomology. 36 (1): 82–93. doi:10.1111/j.1365-2311.2010.01249.x. S2CID 85074319.
- Deventer, S. A.; Herberstein, M. E.; Mayntz, D.; O'Hanlon, J. C.; Schneider, J. M. (2017). "Female fecundity and offspring survival are not increased through sexual cannibalism in the spider Larinioides sclopetarius". Journal of Evolutionary Biology. 30 (12): 2146–2155. doi:10.1111/jeb.13178. ISSN 1420-9101. PMID 28902470.
- Heiling, Astrid M. (1999-06-09). "Why do nocturnal orb-web spiders (Araneidae) search for light?". Behavioral Ecology and Sociobiology. 46 (1): 43–49. doi:10.1007/s002650050590. ISSN 0340-5443. S2CID 25723085.
- Heiling; Herberstein (1999). "The Importance of Being Larger: Intraspecific Competition for Prime Web Sites in Orb-Web Spiders (Araneae, Araneidae)". Behaviour. 136 (5): 669–677. doi:10.1163/156853999501513. ISSN 0005-7959.
- Heiling, Astrid; Herberstein, Marie (1997). Activity patterns in different developmental stages and sexes of Larinioides sclopetarius (Clerck) (Araneae, Araneidae) (PDF). British Arachnological Society. pp. 211–214. ISBN 0-9500093-2-6.
- Herberstein, Marie; Heiling, Astrid (1998). "Does mesh height influence prey length in orb-web spiders (Araneae)?" (PDF). European Journal of Entomology. 95: 367–371.
- Venner, S.; Thevenard, L.; Pasquet, A.; Leborgne, R. (2001-05-01). "Estimation of the Web's Capture Thread Length in Orb-Weaving Spiders: Determining the Most Efficient Formula". Annals of the Entomological Society of America. 94 (3): 490–496. doi:10.1603/0013-8746(2001)094[0490:eotwsc]2.0.co;2. ISSN 0013-8746.
- Herberstein, M.E.; Heiling, A.M. (December 1999). "Asymmetry in spider orb webs: a result of physical constraints?". Animal Behaviour. 58 (6): 1241–1246. doi:10.1006/anbe.1999.1255. ISSN 0003-3472. PMID 10600145. S2CID 24704458.
- Herberstein, M. E.; Heiling, A. M. (October 2001). "Positioning at the hub: does it matter on which side of the web orb-web spiders sit?". Journal of Zoology. 255 (2): 157–163. doi:10.1017/s0952836901001224. ISSN 0952-8369.
- Murakami, Yasuaki (1983-03-01). "Factors determining the prey size of the orb-web spider, Argiope amoena (L. Koch) (Argiopidae)". Oecologia. 57 (1): 72–77. Bibcode:1983Oecol..57...72M. doi:10.1007/BF00379564. ISSN 1432-1939. PMID 28310159. S2CID 22116350.
- "File:Larinioides sclopetarius (Araneidae sp.) female, Arnhem, the Netherlands - 2.jpg" by Bj.schoenmakers is marked under CC0 1.0. To view the terms, visit http://creativecommons.org/publicdomain/zero/1.0/deed.en
- Kralj-Fiser, S.; Sanguino Mostajo, G. A.; Preik, O.; Pekar, S.; Schneider, J. M. (2013-04-18). "Assortative mating by aggressiveness type in orb weaving spiders". Behavioral Ecology. 24 (4): 824–831. doi:10.1093/beheco/art030. hdl:10.1093/beheco/art030. ISSN 1045-2249.
- Kropf, Christian; Bauer, Dina; Schläppi, Thomas; Jacob, Alain (February 2012). "An organic coating keeps orb-weaving spiders (Araneae, Araneoidea, Araneidae) from sticking to their own capture threads: An organic coating protects orb web spiders". Journal of Zoological Systematics and Evolutionary Research. 50 (1): 14–18. doi:10.1111/j.1439-0469.2011.00648.x.
- GUARISCO, HANK (August 2001). "Description of the Egg Sac of Mimetus Notius (Araneae, Mimetidae) and a Case of Egg Predation by Phalacrotophora Epeirae (Diptera, Phoridae)". Journal of Arachnology. 29 (2): 267–269. doi:10.1636/0161-8202(2001)029[0267:doteso]2.0.co;2. ISSN 0161-8202. S2CID 86155933.
- Asis, Josep Daniel; Tormos, Jose; Gayubo, Severiano Fernandez (1996-05-01). "Behavior of Philanthus pulchellus (Hymenoptera: Sphecidae) with a Description of Its Mature Larva". Annals of the Entomological Society of America. 89 (3): 452–458. doi:10.1093/aesa/89.3.452. ISSN 1938-2901.
- Swasko, Mick (July 13, 2012). "Spiders attacking Downtown High-Rises? Not so fast". Chicago Tribune. Retrieved 2021-12-18.
- Laine, Samantha (May 8, 2015). "What if spiders could fly? In Chicago, perhaps they do". Christian Science Monitor. ISSN 0882-7729. Retrieved 2021-12-18.
External links
 Media related to Larinioides sclopetarius at Wikimedia Commons
Media related to Larinioides sclopetarius at Wikimedia Commons- Picture of a L. sclopetarius