Nucleoside-modified messenger RNA
A nucleoside-modified messenger RNA (modRNA) is a synthetic messenger RNA (mRNA) in which some nucleosides are replaced by other naturally modified nucleosides or by synthetic nucleoside analogues.[1] modRNA is used to induce the production of a desired protein in certain cells. An important application is the development of mRNA vaccines, of which the first authorized were COVID-19 vaccines (such as Comirnaty and Spikevax).
Background
mRNA is produced by synthesising a ribonucleic acid (RNA) strand from nucleotide building blocks according to a deoxyribonucleic acid (DNA) template, a process that is called transcription.[2] When the building blocks provided to the RNA polymerase include non-standard nucleosides such as pseudouridine — instead of the standard adenosine, cytidine, guanosine, and uridine nucleosides — the resulting mRNA is described as nucleoside-modified.[3]
Production of protein begins with assembly of ribosomes on the mRNA, the latter then serving as a blueprint for the synthesis of proteins by specifying their amino acid sequence based on the genetic code in the process of protein biosynthesis called translation.[4]
Overview
To induce cells to make proteins that they do not normally produce, it is possible to introduce heterologous mRNA into the cytoplasm of the cell, bypassing the need for transcription. In other words, a blueprint for foreign proteins is "smuggled" into the cells. To achieve this goal, however, one must bypass cellular systems that prevent the penetration and translation of foreign mRNA. There are nearly-ubiquitous enzymes called ribonucleases (also called RNAses) that break down unprotected mRNA.[5] There are also intracellular barriers against foreign mRNA, such as innate immune system receptors, toll-like receptor (TLR) 7 and TLR8, located in endosomal membranes. RNA sensors like TLR7 and TLR8 can dramatically reduce protein synthesis in the cell, trigger release of cytokines such as interferon and TNF-alpha, and when sufficiently intense lead to programmed cell death.[6]
The inflammatory nature of exogenous RNA can be masked by modifying the nucleosides in mRNA.[7] For example, uridine can be replaced with a similar nucleoside such as pseudouridine (Ψ) or N1-methyl-pseudouridine (m1Ψ),[8] and cytosine can be replaced by 5-methylcytosine.[9] Some of these, such as pseudouridine and 5-methylcytosine, occur naturally in eukaryotes,[10] while m1Ψ occurs naturally in archaea.[11] Inclusion of these modified nucleosides alters the secondary structure of the mRNA, which can reduce recognition by the innate immune system while still allowing effective translation.[9]
Significance of untranslated regions
A normal mRNA starts and ends with sections that do not code for amino acids of the actual protein. These sequences at the 5′ and 3′ ends of an mRNA strand are called untranslated regions (UTRs). The two UTRs at their strand ends are essential for the stability of an mRNA and also of a modRNA as well as for the efficiency of translation, i.e. for the amount of protein produced. By selecting suitable UTRs during the synthesis of a modRNA, the production of the target protein in the target cells can be optimised.[5][12]
Delivery
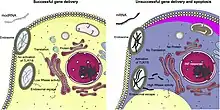
Various difficulties are involved in the introduction of modRNA into certain target cells. First, the modRNA must be protected from ribonucleases.[5] This can be accomplished, for example, by wrapping it in liposomes. Such "packaging" can also help to ensure that the modRNA is absorbed into the target cells. This is useful, for example, when used in vaccines, as nanoparticles are taken up by dendritic cells and macrophages, both of which play an important role in activating the immune system.[13]
Furthermore, it may be desirable that the modRNA applied is introduced into specific body cells. This is the case, for example, if heart muscle cells are to be stimulated to multiply. In this case, the packaged modRNA can be injected directly into an artery such as a coronary artery.[14]
Applications
An important field of application are mRNA vaccines.
Replacing uridine with pseudouridine to evade the innate immune system was pioneered by Karikó and Weissman in 2005.[15][16] They won the 2023 Nobel Prize in Physiology or Medicine as a result of their work.[17]
Another milestone was achieved by demonstrating the life-saving efficacy of nucleoside modified mRNA in a mouse model of a lethal lung disease by the team of Kormann and others in 2011.[18]
N1-methyl-pseudouridine was used in vaccine trials against Zika,[19][20][21] HIV-1,[21] influenza,[21] and Ebola[22] in 2017–2018.[23]: 5
The first authorized for use in humans were COVID-19 vaccines to address SARS-CoV-2.[24][25][26][27][28][29][30] Examples of COVID-19 vaccines using modRNA include those developed by the cooperation of BioNTech/Pfizer (BNT162b2), and by Moderna (mRNA-1273).[31][32][33] The zorecimeran vaccine developed by Curevac, however, uses unmodified RNA,[34] instead relying on codon optimization to minimize the presence of uridine. This vaccine is less effective, however.[35][16]
Other possible uses of modRNA include the regeneration of damaged heart muscle tissue,[36][37] an enzyme-replacement tool[38] and cancer therapy.[39][40]
References
- Chien KR, Zangi L, Lui KO (October 2014). "Synthetic chemically modified mRNA (modRNA): toward a new technology platform for cardiovascular biology and medicine". Cold Spring Harbor Perspectives in Medicine. 5 (1): a014035. doi:10.1101/cshperspect.a014035. PMC 4292072. PMID 25301935.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). From DNA to RNA (4 ed.). Garland Science.
- Pardi N, Weissman D (2017). "Nucleoside Modified mRNA Vaccines for Infectious Diseases". RNA Vaccines. Methods in Molecular Biology. Vol. 1499. Clifton, N.J. pp. 109–121. doi:10.1007/978-1-4939-6481-9_6. ISBN 978-1-4939-6479-6. PMID 27987145.
{{cite book}}: CS1 maint: location missing publisher (link) - Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). The Three Roles of RNA in Protein Synthesis (4th ed.). New York: W. H. Freeman. pp. Sec 4.4.
- Schlake T, Thess A, Fotin-Mleczek M, Kallen KJ (November 2012). "Developing mRNA-vaccine technologies". RNA Biology. 9 (11): 1319–1330. doi:10.4161/rna.22269. PMC 3597572. PMID 23064118.
- Dalpke A, Helm M (June 2012). "RNA mediated Toll-like receptor stimulation in health and disease". RNA Biology. 9 (6): 828–842. doi:10.4161/rna.20206. PMC 3495747. PMID 22617878.
- Karikó K, Buckstein M, Ni H, Weissman D (August 2005). "Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA". Immunity. 23 (2): 165–175. doi:10.1016/j.immuni.2005.06.008. PMID 16111635.
- Andries O, Mc Cafferty S, De Smedt SC, Weiss R, Sanders NN, Kitada T (November 2015). "N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice". Journal of Controlled Release. 217: 337–344. doi:10.1016/j.jconrel.2015.08.051. hdl:1854/LU-6993270. PMID 26342664.
- Svitkin YV, Cheng YM, Chakraborty T, Presnyak V, John M, Sonenberg N (June 2017). "N1-methyl-pseudouridine in mRNA enhances translation through eIF2α-dependent and independent mechanisms by increasing ribosome density". Nucleic Acids Research. 45 (10): 6023–6036. doi:10.1093/nar/gkx135. PMC 5449617. PMID 28334758.
- Hoernes TP, Clementi N, Faserl K, Glasner H, Breuker K, Lindner H, et al. (January 2016). "Nucleotide modifications within bacterial messenger RNAs regulate their translation and are able to rewire the genetic code". Nucleic Acids Research. 44 (2): 852–862. doi:10.1093/nar/gkv1182. PMC 4737146. PMID 26578598.
- Wurm JP, Griese M, Bahr U, Held M, Heckel A, Karas M, et al. (March 2012). "Identification of the enzyme responsible for N1-methylation of pseudouridine 54 in archaeal tRNAs". RNA. 18 (3): 412–420. doi:10.1261/rna.028498.111. PMC 3285930. PMID 22274954.
In contrast, in most archaea this position is occupied by another hypermodified nucleotide: the isosteric N1-methylated pseudouridine.
- Orlandini von Niessen AG, Poleganov MA, Rechner C, Plaschke A, Kranz LM, Fesser S, et al. (April 2019). "Improving mRNA-Based Therapeutic Gene Delivery by Expression-Augmenting 3' UTRs Identified by Cellular Library Screening". Molecular Therapy. 27 (4): 824–836. doi:10.1016/j.ymthe.2018.12.011. PMC 6453560. PMID 30638957.
- Zhao L, Seth A, Wibowo N, Zhao CX, Mitter N, Yu C, Middelberg AP (January 2014). "Nanoparticle vaccines". Vaccine. 32 (3): 327–337. doi:10.1016/j.vaccine.2013.11.069. PMID 24295808.
- Carlsson L, Clarke JC, Yen C, Gregoire F, Albery T, Billger M, et al. (June 2018). "Biocompatible, Purified VEGF-A mRNA Improves Cardiac Function after Intracardiac Injection 1 Week Post-myocardial Infarction in Swine". Molecular Therapy. Methods & Clinical Development. 9: 330–346. doi:10.1016/j.omtm.2018.04.003. PMC 6054703. PMID 30038937.
- Karikó K, Buckstein M, Ni H, Weissman D (August 2005). "Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA". Immunity. 23 (2): 165–175. doi:10.1016/j.immuni.2005.06.008. PMID 16111635.
- Dolgin E (September 2021). "The tangled history of mRNA vaccines". Nature. 597 (7876): 318–324. Bibcode:2021Natur.597..318D. doi:10.1038/d41586-021-02483-w. PMID 34522017. S2CID 237515383.
- https://www.nobelprize.org/prizes/medicine/2023/summary/
- Kormann, Michael S. D.; Hasenpusch, Günther; Aneja, Manish K.; Nica, Gabriela; Flemmer, Andreas W.; Herber-Jonat, Susanne; Huppmann, Marceline; Mays, Lauren E.; Illenyi, Marta; Schams, Andrea; Griese, Matthias; Bittmann, Iris; Handgretinger, Rupert; Hartl, Dominik; Rosenecker, Joseph (February 2011). "Expression of therapeutic proteins after delivery of chemically modified mRNA in mice". Nature Biotechnology. 29 (2): 154–157. doi:10.1038/nbt.1733. ISSN 1546-1696. PMID 21217696. S2CID 205275040.
- Pardi N, Hogan MJ, Pelc RS, Muramatsu H, Andersen H, DeMaso CR, et al. (March 2017). "Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination". Nature. 543 (7644): 248–251. Bibcode:2017Natur.543..248P. doi:10.1038/nature21428. PMC 5344708. PMID 28151488.
we designed a potent anti-ZIKV vaccine … containing the modified nucleoside 1-methylpseudouridine (m1Ψ)
- Richner JM, Himansu S, Dowd KA, Butler SL, Salazar V, Fox JM, et al. (March 2017). "Modified mRNA Vaccines Protect against Zika Virus Infection". Cell. 168 (6): 1114–1125.e10. doi:10.1016/j.cell.2017.02.017. PMC 5388441. PMID 28222903.
The mRNA was synthesized … where the UTP was substituted with 1-methylpseudoUTP
- Pardi N, Hogan MJ, Naradikian MS, Parkhouse K, Cain DW, Jones L, et al. (June 2018). "Nucleoside-modified mRNA vaccines induce potent T follicular helper and germinal center B cell responses". The Journal of Experimental Medicine. 215 (6): 1571–1588. doi:10.1084/jem.20171450. PMC 5987916. PMID 29739835.
In this study, we characterize the immunogenicity of three vaccines consisting of m1Ψ-modified, FPLC-purified mRNA-LNPs encoding HIV-1 envelope (Env), ZIKV prM-E, and influenza virus hemagglutinin (HA)
- Meyer M, Huang E, Yuzhakov O, Ramanathan P, Ciaramella G, Bukreyev A (January 2018). "Modified mRNA-Based Vaccines Elicit Robust Immune Responses and Protect Guinea Pigs From Ebola Virus Disease". The Journal of Infectious Diseases. 217 (3): 451–455. doi:10.1093/infdis/jix592. PMC 5853918. PMID 29281112.
Two mRNA vaccines were synthesized … where the UTP were substituted with 1-methylpseudo UTP
- Morais P, Adachi H, Yu YT (2021-11-04). "The Critical Contribution of Pseudouridine to mRNA COVID-19 Vaccines". Frontiers in Cell and Developmental Biology. 9: 789427. doi:10.3389/fcell.2021.789427. PMC 8600071. PMID 34805188.
- "Pfizer and BioNTech Celebrate Historic First Authorization in the U.S. of Vaccine to Prevent COVID-19". www.businesswire.com. 12 December 2020.
- Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. (December 2020). "Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine". The New England Journal of Medicine. 383 (27): 2603–2615. doi:10.1056/NEJMoa2034577. PMC 7745181. PMID 33301246. S2CID 228087117.
- Hohmann-Jeddi C (2020-11-10). "Hoffnungsträger BNT162b2: Wie funktionieren mRNA-Impfstoffe?". Pharmazeutische Zeitung (in German). Retrieved 2020-11-28.
- Abbasi J (September 2020). "COVID-19 and mRNA Vaccines-First Large Test for a New Approach". JAMA. 324 (12): 1125–1127. doi:10.1001/jama.2020.16866. PMID 32880613. S2CID 221476409.
- Vogel A, Kanevsky I, Che Y, Swanson K, Muik A, Vormehr M, et al. (8 September 2020). "A prefusion SARS-CoV-2 spike RNA vaccine is highly immunogenic and prevents lung infection in non-human primates" (PDF). bioRxiv. doi:10.1101/2020.09.08.280818. S2CID 221589144.
- "Conditions of Authorisation for Pfizer/BioNTech COVID-19 Vaccine" (Decision). Medicines & Healthcare Products Regulatory Agency. 8 December 2020.
- Office of the Commissioner (14 December 2020). "Pfizer-BioNTech COVID-19 Vaccine". www.fda.gov (Decision). US FDA.
- Krammer F (October 2020). "SARS-CoV-2 vaccines in development". Nature. 586 (7830): 516–527. Bibcode:2020Natur.586..516K. doi:10.1038/s41586-020-2798-3. PMID 32967006. S2CID 221887746.
- "Moderna's Pipeline". Moderna. Retrieved 2020-11-28.
- Dolgin E (November 2020). "COVID-19 vaccines poised for launch, but impact on pandemic unclear". Nature Biotechnology: d41587–020–00022-y. doi:10.1038/d41587-020-00022-y. PMID 33239758. S2CID 227176634.
- "COVID-19". CureVac. Retrieved 2020-12-21.
- Dolgin E (June 2021). "CureVac COVID vaccine let-down spotlights mRNA design challenges". Nature. 594 (7864): 483. Bibcode:2021Natur.594..483D. doi:10.1038/d41586-021-01661-0. PMID 34145413. S2CID 235480198.
- Kaur K, Zangi L (December 2020). "Modified mRNA as a Therapeutic Tool for the Heart". Cardiovascular Drugs and Therapy. 34 (6): 871–880. doi:10.1007/s10557-020-07051-4. PMC 7441140. PMID 32822006.
- Zangi L, Lui KO, von Gise A, Ma Q, Ebina W, Ptaszek LM, et al. (October 2013). "Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction". Nature Biotechnology. 31 (10): 898–907. doi:10.1038/nbt.2682. PMC 4058317. PMID 24013197.
- Ter Huurne, Menno; Parker, Benjamin L.; Liu, Ning Qing; Qian, Elizabeth Ling; Vivien, Celine; Karavendzas, Kathy; Mills, Richard J.; Saville, Jennifer T.; Abu-Bonsrah, Dad; Wise, Andrea F.; Hudson, James E.; Talbot, Andrew S.; Finn, Patrick F.; Martini, Paolo G. V.; Fuller, Maria (2023-09-07). "GLA-modified RNA treatment lowers GB3 levels in iPSC-derived cardiomyocytes from Fabry-affected individuals". American Journal of Human Genetics. 110 (9): 1600–1605. doi:10.1016/j.ajhg.2023.07.013. ISSN 1537-6605. PMC 10502840. PMID 37607539.
- McNamara MA, Nair SK, Holl EK (2015). "RNA-Based Vaccines in Cancer Immunotherapy". Journal of Immunology Research. 2015: 794528. doi:10.1155/2015/794528. PMC 4668311. PMID 26665011.
- Verbeke R, Lentacker I, Wayteck L, Breckpot K, Van Bockstal M, Descamps B, et al. (November 2017). "Co-delivery of nucleoside-modified mRNA and TLR agonists for cancer immunotherapy: Restoring the immunogenicity of immunosilent mRNA". Journal of Controlled Release. 266: 287–300. doi:10.1016/j.jconrel.2017.09.041. PMID 28987878. S2CID 20794075.
Further reading
- Badieyan ZS, Evans T (August 2019). "Concise Review: Application of Chemically Modified mRNA in Cell Fate Conversion and Tissue Engineering". Stem Cells Translational Medicine. 8 (8): 833–843. doi:10.1002/sctm.18-0259. PMC 6646692. PMID 30891922.
- Espeseth AS, Cejas PJ, Citron MP, Wang D, DiStefano DJ, Callahan C, et al. (2020). "Modified mRNA/lipid nanoparticle-based vaccines expressing respiratory syncytial virus F protein variants are immunogenic and protective in rodent models of RSV infection". npj Vaccines. 5 (1): 16. doi:10.1038/s41541-020-0163-z. PMC 7021756. PMID 32128257.
Types of nucleic acids | |||||||
|---|---|---|---|---|---|---|---|
| Constituents | |||||||
| Ribonucleic acids (coding, non-coding) |
| ||||||
| Deoxyribonucleic acids | |||||||
| Analogues | |||||||
| Cloning vectors | |||||||
| |||||||