Mossy fiber (hippocampus)
In the hippocampus, the mossy fiber pathway consists of unmyelinated axons projecting from granule cells in the dentate gyrus that terminate on modulatory hilar mossy cells[2][3] and in Cornu Ammonis area 3 (CA3),[4] a region involved in encoding short-term memory.[5][6] These axons were first described as mossy fibers by Santiago Ramón y Cajal as they displayed varicosities along their lengths that gave them a mossy appearance.[7] The axons that make up the pathway emerge from the basal portions of the granule cells and pass through the hilus (or polymorphic cell layer) of the dentate gyrus before entering the stratum lucidum of CA3. Granule cell synapses tend to be glutamatergic (i.e. excitatory), though immunohistological data has indicated that some synapses contain neuropeptidergic elements including opiate peptides such as dynorphin and enkephalin. There is also evidence for co-localization of both GABAergic (i.e. inhibitory) and glutamatergic neurotransmitters within mossy fiber terminals.[8][9] GABAergic and glutamatergic co-localization in mossy fiber boutons has been observed primarily in the developing hippocampus,[10] but in adulthood, evidence suggests that mossy fiber synapses may alternate which neurotransmitter is released through activity-dependent regulation.[11]
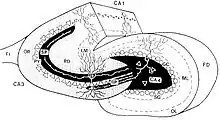
Anatomy
Mossy fibers in the hippocampus project from the dentate gyrus to CA3. The pathway consists of varicose granule cell axons that terminate on the dendrites of hilar mossy cells and pyramidal cells in CA3.[4] They form three morphologically different synaptic terminals, which include large mossy terminals, filopodial extensions within the mossy terminals, and small en passant synaptic varicosities. Each of these synapse types is functionally distinct.[12]
Synaptic terminals
Mossy fibers form multiple synapses with the elaborate dendritic spines of CA3 pyramidal cells in the stratum lucidum of the hippocampus. These complex spines are known as "thorny excrescences."[4][13] Thorny excrescences also cover the proximal dendrites of mossy cells in the hilus. Hilar thorny excrescences are more dense and complex than those in CA3. It has been shown that the axons of granule cells from the dentate gyrus synapse with hilar mossy cells and GABAergic interneurons including basket cells before reaching pyramidal cells in the CA3 region,[4] providing input from the entorhinal cortex through the perforant pathway. Hilar mossy cell activation is thought to be necessary for the proper function of these inhibitory basket cells on CA3 pyramidal cells, although evidence has shown that sodium channel receptors can regulate basket cell function as well.[7]
The three synaptic terminals types – mossy terminals, filopodial extensions, and en passant synaptic varicosities – differ in synaptic output. Large mossy terminals synapse with 11–15 different CA3 pyramidal cells and 7–12 mossy cells.[12] En passant boutons with 25–35 synaptic connections and filopodial extensions with 12–17 make up a significant portion of total granule cell synaptic terminals and are mainly responsible for the excitation of GABAergic interneurons. The type of synaptic terminal expressed therefore dictates the downstream targeting of granule cells.[4][12] The high convergence onto pyramidal cells and divergent projections onto interneurons suggests a primarily modulatory role for the mossy fiber pathway in the hippocampus.[12][2]
The synapses of the mossy fibers contain zinc, which can be stained with a Timm staining.[14]
Projections
The dentate gyrus receives excitatory projections from neurons in layer II of the entorhinal cortex as well as input from surrounding neuroglia.[15] The unmyelinated granule cell axons of the mossy fiber pathway express both GABA receptors and glutamate receptors along their membranes that allow them to be modulated by both excitatory and inhibitory input from nearby glial cells.[16][17] Axons from the entorhinal cortex synapse primarily on the dendritic spines of outer layer dentate granule cells.[18] The entorhinal cortex passes sensory information from neocortical structures to the hippocampal formation.[17] The pathway allows sensory information to reach the hippocampus for encoding.
The mossy fiber pathway itself projects to CA3. Repetitive stimulation of its neurons leads to progressive use-dependent synaptic depression. These short-term changes in plasticity have been shown to be mediated by sodium channels that receive input from neuroglia.[16] The entorhinal cortex also projects directly to CA3, suggesting that the mossy fiber pathway may be functionally similar to the perforant pathway although microcircuits within the dentate gyrus give the mossy fiber pathway a more modulatory role.[19] Projections to the dentate hilus are excitatory by nature and oppose the inhibitory effects of interneurons on hilar mossy cells. The result is an excitatory feedforward loop on mossy cells as a result of activation by the entorhinal cortex.[2][15]
Role in learning and memory
In the mouse, a single mossy fiber projection may make as many as 37 contacts with a single pyramidal cell, but innervates only about a dozen different pyramidal cells. In contrast, a single CA3 pyramidal cell receives input from about 50 different granule cells. It has been shown in rodents that the size of the mossy fiber projections can show large interindividual variations, which are to a large part heritable.[20] In addition, these variations show strong correlations with different types of behavior, mainly, but not exclusively, spatial learning.[21]
See also
References
- Sluyter F, Jamot L, Bertholet JY, Crusio WE (April 2005). "Prenatal exposure to alcohol does not affect radial maze learning and hippocampal mossy fiber sizes in three inbred strains of mouse". Behavioral and Brain Functions. 1 (1): 5. doi:10.1186/1744-9081-1-5. PMC 1143778. PMID 15916699.
- Sun Y, Grieco SF, Holmes TC, Xu X (2017-03-01). "Local and Long-Range Circuit Connections to Hilar Mossy Cells in the Dentate Gyrus". eNeuro. 4 (2): ENEURO.0097–17.2017. doi:10.1523/ENEURO.0097-17.2017. PMC 5396130. PMID 28451637.
- Henze DA, Buzsáki G (2007). "Hilar mossy cells: functional identification and activity in vivo". The Dentate Gyrus: A Comprehensive Guide to Structure, Function, and Clinical Implications. Progress in Brain Research. Vol. 163. pp. 199–216. doi:10.1016/s0079-6123(07)63012-x. ISBN 9780444530158. PMID 17765720.
- Amaral DG, Scharfman HE, Lavenex P (2007). "The dentate gyrus: fundamental neuroanatomical organization (dentate gyrus for dummies)". The Dentate Gyrus: A Comprehensive Guide to Structure, Function, and Clinical Implications. Progress in Brain Research. Vol. 163. pp. 3–22. doi:10.1016/S0079-6123(07)63001-5. ISBN 9780444530158. PMC 2492885. PMID 17765709.
- Kesner RP (November 2007). "Behavioral functions of the CA3 subregion of the hippocampus" (PDF). Learning & Memory. 14 (11): 771–81. doi:10.1101/lm.688207. PMID 18007020.
- Farovik A, Dupont LM, Eichenbaum H (January 2010). "Distinct roles for dorsal CA3 and CA1 in memory for sequential nonspatial events". Learning & Memory. 17 (1): 12–17. doi:10.1101/lm.1616209. PMC 2807176. PMID 20028733.
- Scharfman HE, Myers CE (2012). "Hilar mossy cells of the dentate gyrus: a historical perspective". Frontiers in Neural Circuits. 6: 106. doi:10.3389/fncir.2012.00106. PMC 3572871. PMID 23420672.
- Sandler R, Smith AD (January 1991). "Coexistence of GABA and glutamate in mossy fiber terminals of the primate hippocampus: an ultrastructural study". The Journal of Comparative Neurology. 303 (2): 177–92. doi:10.1002/cne.903030202. PMID 1672874. S2CID 42502573.
- Gutiérrez R, Romo-Parra H, Maqueda J, Vivar C, Ramìrez M, Morales MA, Lamas M (July 2003). "Plasticity of the GABAergic phenotype of the "glutamatergic" granule cells of the rat dentate gyrus". The Journal of Neuroscience. 23 (13): 5594–8. doi:10.1523/jneurosci.23-13-05594.2003. PMC 6741238. PMID 12843261.
- Safiulina VF, Fattorini G, Conti F, Cherubini E (January 2006). "GABAergic signaling at mossy fiber synapses in neonatal rat hippocampus". The Journal of Neuroscience. 26 (2): 597–608. doi:10.1523/JNEUROSCI.4493-05.2006. PMC 6674413. PMID 16407558.
- Caiati MD (January 2013). "Is GABA co-released with glutamate from hippocampal mossy fiber terminals?". The Journal of Neuroscience. 33 (5): 1755–6. doi:10.1523/JNEUROSCI.5019-12.2013. PMC 6619118. PMID 23365214.
- Acsády L, Kamondi A, Sík A, Freund T, Buzsáki G (May 1998). "GABAergic cells are the major postsynaptic targets of mossy fibers in the rat hippocampus". The Journal of Neuroscience. 18 (9): 3386–403. doi:10.1523/jneurosci.18-09-03386.1998. PMC 6792657. PMID 9547246.
- Gonzales RB, DeLeon Galvan CJ, Rangel YM, Claiborne BJ (February 2001). "Distribution of thorny excrescences on CA3 pyramidal neurons in the rat hippocampus". The Journal of Comparative Neurology. 430 (3): 357–68. doi:10.1002/1096-9861(20010212)430:3<357::aid-cne1036>3.0.co;2-k. PMID 11169473.
- Danscher G, Zimmer J (February 1978). "An improved Timm sulphide silver method for light and electron microscopic localization of heavy metals in biological tissues". Histochemistry. 55 (1): 27–40. doi:10.1007/bf00496691. PMID 76622. S2CID 2771576.
- Jonas P, Lisman J (2014). "Structure, function, and plasticity of hippocampal dentate gyrus microcircuits". Frontiers in Neural Circuits. 8: 107. doi:10.3389/fncir.2014.00107. PMC 4159971. PMID 25309334.
- Ohura S, Kamiya H (January 2018). "Short-Term Depression of Axonal Spikes at the Mouse Hippocampal Mossy Fibers and Sodium Channel-Dependent Modulation". eNeuro. 5 (1): ENEURO.0415–17.2018. doi:10.1523/ENEURO.0415-17.2018. PMC 5820996. PMID 29468192.
- Amaral DG, Kondo H, Lavenex P (May 2014). "An analysis of entorhinal cortex projections to the dentate gyrus, hippocampus, and subiculum of the neonatal macaque monkey". The Journal of Comparative Neurology. 522 (7): 1485–505. doi:10.1002/cne.23469. PMC 4384686. PMID 24122645.
- Witter MP (2007). "The perforant path: projections from the entorhinal cortex to the dentate gyrus". The Dentate Gyrus: A Comprehensive Guide to Structure, Function, and Clinical Implications. Progress in Brain Research. Vol. 163. pp. 43–61. doi:10.1016/S0079-6123(07)63003-9. ISBN 9780444530158. PMID 17765711.
- Henze DA, Urban NN, Barrionuevo G (2000). "The multifarious hippocampal mossy fiber pathway: a review". Neuroscience. 98 (3): 407–27. doi:10.1016/s0306-4522(00)00146-9. PMID 10869836. S2CID 11663807.
- Crusio WE, Genthner-Grimm G, Schwegler H (2007). "A quantitative-genetic analysis of hippocampal variation in the mouse". Journal of Neurogenetics. 21 (4): 197–208. doi:10.1080/01677060701715827. PMID 18161583. S2CID 7334116.
- Crusio WE, Schwegler H (April 2005). "Learning spatial orientation tasks in the radial-maze and structural variation in the hippocampus in inbred mice". Behavioral and Brain Functions. 1 (1): 3. doi:10.1186/1744-9081-1-3. PMC 1143776. PMID 15916698.