Multicopy single-stranded DNA
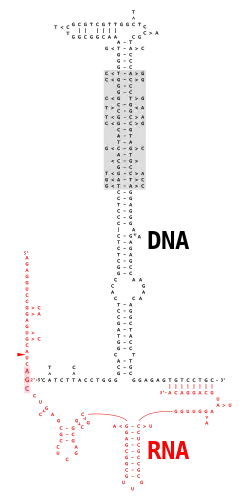
Multicopy single-stranded DNA (msDNA) is a type of extrachromosomal satellite DNA that consists of a single-stranded DNA molecule covalently linked via a 2'-5'phosphodiester bond to an internal guanosine of an RNA molecule. The resultant DNA/RNA chimera possesses two stem-loops joined by a branch similar to the branches found in RNA splicing intermediates. The coding region for msDNA, called a "retron", also encodes a type of reverse transcriptase, which is essential for msDNA synthesis.[2]
Discovery
Before the discovery of msDNA in myxobacteria,[3][4] a group of swarming, soil-dwelling bacteria, it was thought that the enzymes known as reverse transcriptases (RT) existed only in eukaryotes and viruses. The discovery led to an increase in research of the area. As a result, msDNA has been found to be widely distributed among bacteria, including various strains of Escherichia coli and pathogenic bacteria.[5] Further research discovered similarities between HIV-encoded reverse transcriptase and an open reading frame (ORF) found in the msDNA coding region. Tests confirmed the presence of reverse transcriptase activity in crude lysates of retron-containing strains.[6] Although an RNase H domain was tentatively identified in the retron ORF, it was later found that the RNase H activity required for msDNA synthesis is actually supplied by the host.[7]
Retrons
The discovery of msDNA has led to broader questions regarding where reverse transcriptase originated, as genes encoding for reverse transcriptase (not necessarily associated with msDNA) have been found in prokaryotes, eukaryotes, viruses and even archaea. After a DNA fragment coding for the production of msDNA in E. coli was discovered,[8] it was conjectured that bacteriophages might have been responsible for the introduction of the RT gene into E. coli.[9] These discoveries suggest that reverse transcriptase played a role in the evolution of viruses from bacteria, with one hypothesis stating that, with the help of reverse transcriptase, viruses may have arisen as a breakaway msDNA gene that acquired a protein coat. Since nearly all RT genes function in retrovirus replication and/or the movement of transposable elements, it is reasonable to imagine that retrons might be mobile genetic elements, but there has been little supporting evidence for such a hypothesis, save for the observed fact that msDNA is widely yet sporadically dispersed among bacterial species in a manner suggestive of both horizontal and vertical transfer.[5][10][11] Since it is not known whether retron sequences per se represent mobile elements, retrons are functionally defined by their ability to produce msDNA while deliberately avoiding speculation about other possible activities.
Function
The function of msDNA remains unknown even though many copies are present within cells. Knockout mutations that do not express msDNA are viable, so the production of msDNA is not essential to life under laboratory conditions. Over-expression of msDNA is mutagenic, apparently as a result of titrating out repair proteins by the mismatched base pairs that are typical of their structure.[10] It has been suggested that msDNA may have some role in pathogenicity or the adaptation to stressful conditions.[12] Sequence comparison of msDNAs from Myxococcus xanthus, Stigmatella aurantiaca,[1] and many other bacteria[5][12] reveal conserved and hypervariable domains reminiscent of conserved and hypervariable sequences found in allorecognition molecules.[13] The major msDNAs of M. xanthus and S. aurantiaca, for instance, share 94% sequence homology except within a 19 base-pair domain that shares sequence homology of only 42%.[1] The presence of such domains is significant because myxobacteria exhibit complex cooperative social behaviors including swarming and formation of fruiting bodies, while E. coli and other pathogenic bacteria form biofilms that exhibit enhanced antibiotic and detergent resistance. The sustainability of social assemblies that require significant individual investment of energy is generally dependent on the evolution of allorecognition mechanisms that enable groups to distinguish self versus non-self.[14]
Biosynthesis
Biosynthesis of msDNA is purported to follow a unique pathway found nowhere else in DNA/RNA biochemistry. Because of the similarity of the 2'-5' branch junction to the branch junctions found in RNA splicing intermediates, it might at first have been expected that branch formation would be via spliceosome- or ribozyme-mediated ligation. Surprisingly, however, experiments in cell-free systems using purified retron reverse transcriptase indicate that cDNA synthesis is directly primed from the 2'-OH group of the specific internal G residue of the primer RNA.[15] The RT recognizes specific stem-loop structures in the precursor RNA, rendering synthesis of msDNA by the RT highly specific to its own retron.[16] The priming of msDNA synthesis offers a fascinating challenge to our understanding of DNA synthesis. DNA polymerases (which include RT) share highly conserved structural features, which means that their active catalytic sites vary little from species to species, or even between DNA polymerases using DNA as a template, versus DNA polymerases using RNA as a template. The catalytic region of eukaryotic reverse transcriptase comprises three domains termed the "fingers", "palm", and "thumb" which hold the double-stranded primer-template in a right-hand grip with the 3'-OH of the primer buried in the active site of the polymerase,[17] a cluster of highly conserved acidic and polar residues situated on the palm between what would be the index and middle fingers. In eukaryotic RTs, the RNase H domain lies on the wrist below the base of the thumb, but retron RTs lack RNase H activity. The nucleic acid binding cleft, extending from the polymerase active site to the RNase H active site, is about 60 Å in length in eukaryotic RTs, corresponding to nearly two helical turns.[18] When eukaryotic RT extends a conventional primer, the growing DNA/RNA double helix spirals along the cleft, and as the double helix passes the RNase H domain, the template RNA is digested to release the nascent strand of cDNA. In the case of msDNA primer extension, however, a long strand of RNA remains attached to the 3'-OH of the priming G. Although it is possible to model an RT-primer template complex which would make the 2'-OH accessible for the priming reaction,[16] further extension of the DNA strand presents a problem: as DNA synthesis progresses, the bulky RNA strand extending from the 3'-OH needs somehow to spiral down the binding cleft without being blocked by steric hindrance. To overcome this issue, the msDNA reverse transcriptase clearly would require special features not shared by other RTs.[10]
References
- Dhundale A, Lampson B, Furuichi T, Inouye M, Inouye S (December 1987). "Structure of msDNA from Myxococcus xanthus: evidence for a long, self-annealing RNA precursor for the covalently linked, branched RNA". Cell. 51 (6): 1105–12. doi:10.1016/0092-8674(87)90596-4. PMID 2446773. S2CID 21762469.
- Inouye S, Herzer PJ, Inouye M (February 1990). "Two independent retrons with highly diverse reverse transcriptases in Myxococcus xanthus". Proceedings of the National Academy of Sciences of the United States of America. 87 (3): 942–5. Bibcode:1990PNAS...87..942I. doi:10.1073/pnas.87.3.942. PMC 53385. PMID 1689062.
- Yee T, Furuichi T, Inouye S, Inouye M (August 1984). "Multicopy single-stranded DNA isolated from a gram-negative bacterium, Myxococcus xanthus". Cell. 38 (1): 203–9. doi:10.1016/0092-8674(84)90541-5. PMID 6088065. S2CID 41165293.
- Furuichi T, Inouye S, Inouye M (January 1987). "Biosynthesis and structure of stable branched RNA covalently linked to the 5' end of multicopy single-stranded DNA of Stigmatella aurantiaca". Cell. 48 (1): 55–62. doi:10.1016/0092-8674(87)90355-2. PMID 2431795. S2CID 32376617.
- Das R, Shimamoto T, Hosen SM, Arifuzzaman M (2011). "Comparative Study of different msDNA (multicopy single-stranded DNA) structures and phylogenetic comparison of reverse transcriptases (RTs): evidence for vertical inheritance" (PDF). Bioinformation. 7 (4): 176–9. doi:10.6026/97320630007176. PMC 3218519. PMID 22102774.
- Lampson BC, Sun J, Hsu MY, Vallejo-Ramirez J, Inouye S, Inouye M (February 1989). "Reverse transcriptase in a clinical strain of Escherichia coli: production of branched RNA-linked msDNA" (PDF). Science. 243 (4894 Pt 1): 1033–8. Bibcode:1989Sci...243.1033L. doi:10.1126/science.2466332. PMID 2466332. Archived from the original (PDF) on 2014-12-22. Retrieved 2012-02-08.
- Lima TM, Lim D (May 1995). "Isolation and characterization of host mutants defective in msDNA synthesis: role of ribonuclease H in msDNA synthesis". Plasmid. 33 (3): 235–8. doi:10.1006/plas.1995.1026. PMID 7568472.
- Hsu MY, Inouye M, Inouye S (December 1990). "Retron for the 67-base multicopy single-stranded DNA from Escherichia coli: a potential transposable element encoding both reverse transcriptase and Dam methylase functions". Proceedings of the National Academy of Sciences of the United States of America. 87 (23): 9454–8. Bibcode:1990PNAS...87.9454H. doi:10.1073/pnas.87.23.9454. PMC 55184. PMID 1701261.
- Inouye S.; Inouye M. (1993). "Bacterial Reverse Transcriptase". In Goff, Stephen and Anna M. Skalka (ed.). Reverse transcriptase. Cold Spring Harbor monograph series. Vol. 23. Plainview, N.Y: Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-382-4.
- Lampson BC, Inouye M, Inouye S (2005). "Retrons, msDNA, and the bacterial genome" (PDF). Cytogenetic and Genome Research. 110 (1–4): 491–9. doi:10.1159/000084982. PMID 16093702. S2CID 24854188. Archived from the original (PDF) on 2016-03-05. Retrieved 2012-02-08.
- Simon DM, Zimmerly S (December 2008). "A diversity of uncharacterized reverse transcriptases in bacteria". Nucleic Acids Research. 36 (22): 7219–29. doi:10.1093/nar/gkn867. PMC 2602772. PMID 19004871.
- Das R, Shimamoto T, Arifuzzaman M (2011). "A Novel msDNA (Multicopy Single-Stranded DNA) Strain Present in Yersinia frederiksenii ATCC 33641 Contig01029 Enteropathogenic Bacteria with the Genomic Analysis of It's Retron". Journal of Pathogens. 2011 (693769): 693769. doi:10.4061/2011/693769. PMC 3335539. PMID 22567337.
- Sherman LA, Chattopadhyay S (1993). "The molecular basis of allorecognition". Annual Review of Immunology. 11: 385–402. doi:10.1146/annurev.iy.11.040193.002125. PMID 8476567.
- Buss, Leo (2006). The Evolution of Individuality. Princeton University Press. ISBN 978-0-691-08469-5.
- Shimamoto T, Kawanishi H, Tsuchiya T, Inouye S, Inouye M (June 1998). "In vitro synthesis of multicopy single-stranded DNA, using separate primer and template RNAs, by Escherichia coli reverse transcriptase". Journal of Bacteriology. 180 (11): 2999–3002. doi:10.1128/JB.180.11.2999-3002.1998. PMC 107272. PMID 9603895.
- Inouye S, Hsu MY, Xu A, Inouye M (October 1999). "Highly specific recognition of primer RNA structures for 2'-OH priming reaction by bacterial reverse transcriptases". The Journal of Biological Chemistry. 274 (44): 31236–44. doi:10.1074/jbc.274.44.31236. PMID 10531319.
- Jacobo-Molina A, Ding J, Nanni RG, Clark AD, Lu X, Tantillo C, Williams RL, Kamer G, Ferris AL, Clark P (July 1993). "Crystal structure of human immunodeficiency virus type 1 reverse transcriptase complexed with double-stranded DNA at 3.0 A resolution shows bent DNA". Proceedings of the National Academy of Sciences of the United States of America. 90 (13): 6320–4. Bibcode:1993PNAS...90.6320J. doi:10.1073/pnas.90.13.6320. PMC 46920. PMID 7687065.
- Sarafianos SG, Das K, Tantillo C, Clark AD, Ding J, Whitcomb JM, Boyer PL, Hughes SH, Arnold E (March 2001). "Crystal structure of HIV-1 reverse transcriptase in complex with a polypurine tract RNA:DNA". The EMBO Journal. 20 (6): 1449–61. doi:10.1093/emboj/20.6.1449. PMC 145536. PMID 11250910.
Further reading
- Lampson B, Inouye M, Inouye S (2001). The msDNAs of bacteria. Progress in Nucleic Acid Research and Molecular Biology. Vol. 67. pp. 65–91. doi:10.1016/S0079-6603(01)67025-9. ISBN 9780125400671. PMID 11525386.
- Zimmerly, Steven (2005). "Mobile introns and retroelements in bacteria". In Mullany, Peter (ed.). The Dynamic Bacterial Genome. Advances in Molecular and Cellular Microbiology. Vol. 8. Cambridge University Press. pp. 121–148. doi:10.1017/CBO9780511541544.004. ISBN 978-0-511-54154-4.
Types of nucleic acids | |||||||
|---|---|---|---|---|---|---|---|
| Constituents | |||||||
| Ribonucleic acids (coding, non-coding) |
| ||||||
| Deoxyribonucleic acids | |||||||
| Analogues | |||||||
| Cloning vectors | |||||||
| |||||||