Perilipin-1
Perilipin, also known as lipid droplet-associated protein, perilipin 1, or PLIN, is a protein that, in humans, is encoded by the PLIN gene.[5] The perilipins are a family of proteins that associate with the surface of lipid droplets. Phosphorylation of perilipin is essential for the mobilization of fats in adipose tissue.[6]
| PLIN1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | PLIN1, FPLD4, PERI, PLIN, perilipin 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 170290 MGI: 1890505 HomoloGene: 2001 GeneCards: PLIN1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Perilipin family of proteins
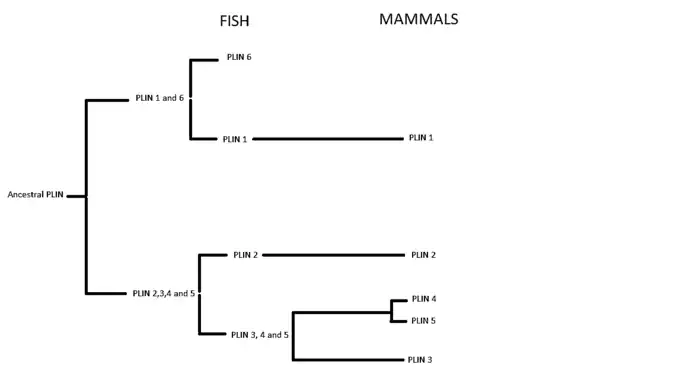
Perilipin is part of a gene family with six currently-known members. In vertebrates, closely related genes include adipophilin (also known as adipose differentiation-related protein or Perilipin 2), TIP47 (Perilipin 3), Perilipin 4 and Perilipin 5 (also called MLDP, LSDP5, or OXPAT). Insects express related proteins, LSD1 and LSD2, in fat bodies.[7] The yeast Saccharomyces cerevisiae expresses PLN1 (formerly PET10), that stabilizes lipid droplets and aids in their assembly.[8]
Evolution
The perilipins are considered to have their origins in a common ancestral gene which, during the first and second vertebrate genome duplication, gave rise to six types of PLIN genes.[9]
Composition and structure
Human perilipin
Human perilipin-1 is composed by 522 amino acids, which add up to a molecular mass of 55.990 kDa. It presents an estimated number of 15 phosphorylation sites (residues 81, 85, 126, 130, 132, 137, 174, 299, 301, 382, 384, 408, 436, 497, 499 and 522)[11] from which 3 -those in bold- have been suggested to be relevant for stimulated-lipolysis through PKA phosphorylation - they correspond respectively to PKA Phosphorylation sites 1, 5 and 6.[12] A compositional bias of Glutamic acid can be found between residues 307 and 316.[13] Its secondary structure has been suggested to be conformed exclusively by partially hydrophobic α-helixes,[10] as well as the respective coils and bends.
Whereas perilipin-1 is coded by a single gene, alternative mRNA splicing processes can lead to three protein isoforms (Perilipin A, B and C). Both Perilipin A and B present common N-terminal regions, differing in the C-terminal ones.[14] Concretely, beginning from the N-terminal of Perilipin-1, a PAT domain—characteristic of its protein family—can be found, followed by an also characteristic repeated sequence of 13 residues which form amphipathic helixes with an active role in linking membranes[15] and a 4-helix bundle before the C-terminal carbon.[16] In Perilipin A, lipophile nature is conferred by the slightly hydrophobic amino acids concentrated in the central 25% of the sequence, region that anchors the protein to the core of the lipid droplet.[17]
| Perilipin | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||
| Symbol | Perilipin | ||||||||||
| Pfam | PF03036 | ||||||||||
| InterPro | IPR004279 | ||||||||||
| |||||||||||
Murine perilipin
Unlike its human ortholog, murine perilipin is composed of 517 amino acids in the primary structure of which several regions can be identified. Three moderately hydrophobic sequences (H1, H2, H3) of 18 rem (243-260 aa), 23 rem (320-332 aa) and 16 rem (349-364 aa) can be identified in the centre of the protein, as well as an acidic region of 28 residues where both glutamic and aspartic acids add up to 19 of them. Five sequences 18 residues long that could form amphipathic β-pleated sheets—according to a prediction made through LOCATE program—are found between aa 111 and 182. Serines occupying positions 81, 222, 276, 433, 492 and 517 act as phosphorylation sites -numbered from 1 to 6- for PKA,[18] as well as several other threonines and serines which add up to 27 phosphorylation sites.[19]
Function
Perilipin is a protein that coats lipid droplets (LDs) in adipocytes,[20] the fat-storing cells in adipose tissue. In fact, PLIN1 is greatly expressed in white adipocytes.[21]
It controls adipocyte lipid metabolism.[22] It handles essential functions in the regulation of basal and hormonally stimulated lipolysis[23] and also rises the formation of large LDs which implies an increase in the synthesis of triglycerides.[21]
In humans, Perilipin A is the most abundant protein associated with the adipocyte LDs[7] and lower PLIN1 expression is related with higher rates of lipolysis.[24]
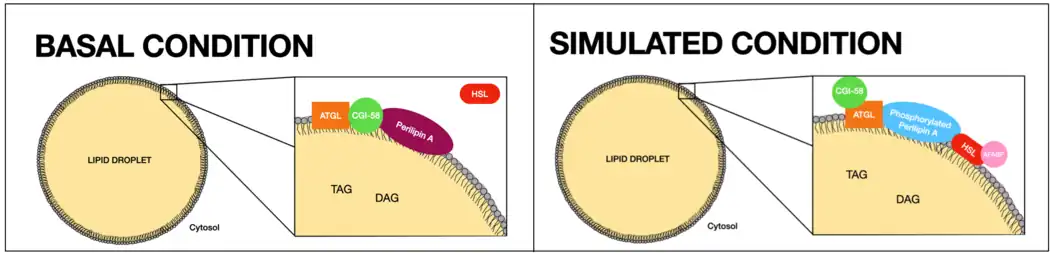
Under basal conditions, Perilipin acts as a protective coating of LDs from the body's natural lipases, such as hormone-sensitive lipase (HSL) and adipose triglyceride lipase (ATGL),[25][24] which break triglycerides into glycerol and free fatty acids for use in lipid metabolism.[6]
In times of energy deficit, Perilipin is hyperphosphorylated by PKA following β-adrenergic receptor activation.[6] Phosphorylated perilipin changes conformation, exposing the stored lipids to hormone-sensitive lipase-mediated lipolysis.
Modulator of adipocyte lipid metabolism
Specifically, in the basal state Perilipin A allows a low level of basal lipolysis[26] by reducing the access of cytosolic lipases to stored triacylglycerol in LDs.[23] It is found at their surface in a complex with CGI-58, the co-activator of ATGL. ATGL might also be in this complex but it is quiescent.[27]
Under lipolytically stimulated conditions, PKA is activated and phosphorylates up to 6 Serine residues on Perilipin A (Ser81, 222, 276, 433, 492, and 517) and 2 on HSL (Ser659, and 660).[27] Although PKA also phosphorylates HSL, which can increase its activity, the more than 50-fold increase in fat mobilization (triggered by epinephrine) is primarily due to Perilipin phosphorylation.
Then, Phosphorylated HSL translocates to the LD surface and associates with Perilipin A and Adipocyte fatty acid-binding protein (AFABP).[27] Consequently, HSL gains access to triacylglycerol (TAG) and diacylglycerol (DAG), substrates in LDs. Also, CGI-58 separates from the LD outer layer which leads to a redistribution of ATGL.[23] In particular, ATGL interacts with Perilipin A through phosphorylated Ser517.[27]
As a result, PKA phosphorylation implies an enriched colocation of HLS and ATGL which facilitates maximal lipolysis by the two lipases.[23]
Clinical significance
Perilipin is an important regulator of lipid storage.[6] Both an overexpression or deficiency of the protein, caused by a mutation, lead to severe health issues.
Overexpression
Perilipin expression is elevated in obese animals and humans. Polymorphisms in the human perilipin (PLIN) gene have been associated with variance in body-weight regulation and may be a genetic influence on obesity risk in humans.[28]
This protein can be modified by O-linked acetylglucosamine (O-GlNac) moieties and the enzyme that intervenes is O-GlcNAc transferase (OGT). An abundance of OGT obstructs lipolysis and benefits diet-induced obesity and whole-body insulin resistance. Studies also propose that an overexpression of adipose O-GlcNAc signaling is a molecular expression of obesity and diabetes in humans.[29]
Deficiency
Perilipin-null mice eat more food than wild-type mice, but gain 1/3 less fat than wild-type mice on the same diet; perilipin-null mice are thinner, with more lean muscle mass.[30] Perilipin-null mice also exhibit enhanced leptin production and a greater tendency to develop insulin resistance than wild-type mice. Even though perilipin-null mice present less fat mass and a higher insulin resistance, they do not show signs of a whole lipodystrophic phenotype.[31]
In humans, studies suggest that a deficiency of PLIN1 causes lipodystrophic syndromes,[32] which disables the optimal accumulation of triglycerides in adipocytes that derives in an abnormal deposition of lipids in tissues such as skeletal muscle and liver. The storage of lipids in the liver leads to insulin resistance and hypertriglyceridemia. Affected patients are characterized by a subcutaneous fat with smaller than normal adipocytes, macrophage infiltration and fibrosis.
These findings affirm a new primary form of inherited lipodystrophy and emphasize on the severe metabolic consequences of a defect in the formation of lipid droplets in adipose tissue.
In particular, variants 13041A>G and 14995A>T have been associated with increased risk of obesity in women and 11482G>A has been associated with decreased perilipin expression and increased lipolysis in women.[33][34]
References
- GRCh38: Ensembl release 89: ENSG00000166819 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000030546 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: PLIN perilipin".
- Mobilization and Cellular Uptake of Stored Fats (with Animation)
- Brasaemle DL, Subramanian V, Garcia A, Marcinkiewicz A, Rothenberg A (June 2009). "Perilipin A and the control of triacylglycerol metabolism". Molecular and Cellular Biochemistry. 326 (1–2): 15–21. doi:10.1007/s11010-008-9998-8. PMID 19116774. S2CID 19802945.
- Gao Q, Binns DD, Kinch LN, Grishin NV, Ortiz N, Chen X, Goodman JM (October 2017). "Pet10p is a yeast perilipin that stabilizes lipid droplets and promotes their assembly". The Journal of Cell Biology. 216 (10): 3199–3217. doi:10.1083/jcb.201610013. PMC 5626530. PMID 28801319.
- "Perilipin-5", Wikipedia, 2020-11-03, retrieved 2020-11-09
- Noureldein MH (2014). "In silico discovery of a perilipin 1 inhibitor to be used as a new treatment for obesity". European Review for Medical and Pharmacological Sciences. 18 (4): 457–60. PMID 24610610.
- Bian Y, Song C, Cheng K, Dong M, Wang F, Huang J, et al. (January 2014). "An enzyme assisted RP-RPLC approach for in-depth analysis of human liver phosphoproteome". Journal of Proteomics. 96: 253–62. doi:10.1016/j.jprot.2013.11.014. PMID 24275569.
- Sztalryd C, Xu G, Dorward H, Tansey JT, Contreras JA, Kimmel AR, Londos C (June 2003). "Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation". The Journal of Cell Biology. 161 (6): 1093–103. doi:10.1083/jcb.200210169. PMC 2172984. PMID 12810697.
- "PLIN1 - Perilipin-1 - Homo sapiens (Human) - PLIN1 gene & protein". www.uniprot.org. Retrieved 2020-11-01.
- Londos C, Brasaemle DL, Schultz CJ, Segrest JP, Kimmel AR (February 1999). "Perilipins, ADRP, and other proteins that associate with intracellular neutral lipid droplets in animal cells". Seminars in Cell & Developmental Biology. 10 (1): 51–8. doi:10.1006/scdb.1998.0275. PMID 10355028.
- Rowe ER, Mimmack ML, Barbosa AD, Haider A, Isaac I, Ouberai MM, et al. (March 2016). "Conserved Amphipathic Helices Mediate Lipid Droplet Targeting of Perilipins 1-3". The Journal of Biological Chemistry. 291 (13): 6664–78. doi:10.1074/jbc.M115.691048. PMC 4807253. PMID 26742848.
- Itabe H, Yamaguchi T, Nimura S, Sasabe N (April 2017). "Perilipins: a diversity of intracellular lipid droplet proteins". Lipids in Health and Disease. 16 (1): 83. doi:10.1186/s12944-017-0473-y. PMC 5410086. PMID 28454542.
- Garcia A, Sekowski A, Subramanian V, Brasaemle DL (January 2003). "The central domain is required to target and anchor perilipin A to lipid droplets". The Journal of Biological Chemistry. 278 (1): 625–35. doi:10.1074/jbc.M206602200. PMID 12407111. S2CID 12795601.
- Zhang HH, Souza SC, Muliro KV, Kraemer FB, Obin MS, Greenberg AS (December 2003). "Lipase-selective functional domains of perilipin A differentially regulate constitutive and protein kinase A-stimulated lipolysis". The Journal of Biological Chemistry. 278 (51): 51535–42. doi:10.1074/jbc.M309591200. PMID 14527948. S2CID 8227051.
- Rogne M, Chu DT, Küntziger TM, Mylonakou MN, Collas P, Tasken K (June 2018). Parton RG (ed.). "OPA1-anchored PKA phosphorylates perilipin 1 on S522 and S497 in adipocytes differentiated from human adipose stem cells". Molecular Biology of the Cell. 29 (12): 1487–1501. doi:10.1091/mbc.E17-09-0538. PMC 6014102. PMID 29688805.
- Greenberg AS, Egan JJ, Wek SA, Garty NB, Blanchette-Mackie EJ, Londos C (June 1991). "Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets". The Journal of Biological Chemistry. 266 (17): 11341–6. doi:10.1016/S0021-9258(18)99168-4. PMID 2040638.
- Shijun L, Khan R, Raza SH, Jieyun H, Chugang M, Kaster N, et al. (May 2020). "Function and characterization of the promoter region of perilipin 1 (PLIN1): Roles of E2F1, PLAG1, C/EBPβ, and SMAD3 in bovine adipocytes". Genomics. 112 (3): 2400–2409. doi:10.1016/j.ygeno.2020.01.012. PMID 31981700. S2CID 210912743.
- "UniProtKB - O60240 (PLIN1_HUMAN)".
- Brasaemle DL (December 2007). "Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis". Journal of Lipid Research. 48 (12): 2547–59. doi:10.1194/jlr.R700014-JLR200. PMID 17878492. S2CID 38744670.
- Grahn TH, Zhang Y, Lee MJ, Sommer AG, Mostoslavsky G, Fried SK, et al. (March 2013). "FSP27 and PLIN1 interaction promotes the formation of large lipid droplets in human adipocytes". Biochemical and Biophysical Research Communications. 432 (2): 296–301. doi:10.1016/j.bbrc.2013.01.113. PMC 3595328. PMID 23399566.
- Wong K (2000-11-29). "Making Fat-proof Mice". Scientific American. Retrieved 2009-05-22.
- Sztalryd C, Brasaemle DL (October 2017). "The perilipin family of lipid droplet proteins: Gatekeepers of intracellular lipolysis". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1862 (10 Pt B): 1221–1232. doi:10.1016/j.bbalip.2017.07.009. PMC 5595658. PMID 28754637.
- Bickel PE, Tansey JT, Welte MA (June 2009). "PAT proteins, an ancient family of lipid droplet proteins that regulate cellular lipid stores". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1791 (6): 419–40. doi:10.1016/j.bbalip.2009.04.002. PMC 2782626. PMID 19375517.
- Soenen S, Mariman EC, Vogels N, Bouwman FG, den Hoed M, Brown L, Westerterp-Plantenga MS (March 2009). "Relationship between perilipin gene polymorphisms and body weight and body composition during weight loss and weight maintenance". Physiology & Behavior. 96 (4–5): 723–8. doi:10.1016/j.physbeh.2009.01.011. PMID 19385027. S2CID 24747708.
- Yang Y, Fu M, Li MD, Zhang K, Zhang B, Wang S, et al. (January 2020). "O-GlcNAc transferase inhibits visceral fat lipolysis and promotes diet-induced obesity". Nature Communications. 11 (1): 181. Bibcode:2020NatCo..11..181Y. doi:10.1038/s41467-019-13914-8. PMC 6954210. PMID 31924761.
- telegraph.co.uk, 19 June 2001, Highfield R (2000-11-29). "Couch potato mice discover the lazy way to stay slim". The Daily Telegraph. London. Retrieved 2008-09-03.
- Tansey JT, Sztalryd C, Gruia-Gray J, Roush DL, Zee JV, Gavrilova O, et al. (May 2001). "Perilipin ablation results in a lean mouse with aberrant adipocyte lipolysis, enhanced leptin production, and resistance to diet-induced obesity". Proceedings of the National Academy of Sciences of the United States of America. 98 (11): 6494–9. Bibcode:2001PNAS...98.6494T. doi:10.1073/pnas.101042998. PMC 33496. PMID 11371650.
- Gandotra S, Le Dour C, Bottomley W, Cervera P, Giral P, Reznik Y, et al. (February 2011). "Perilipin deficiency and autosomal dominant partial lipodystrophy". The New England Journal of Medicine. 364 (8): 740–8. doi:10.1056/NEJMoa1007487. PMC 3773916. PMID 21345103.
- Qi L, Shen H, Larson I, Schaefer EJ, Greenberg AS, Tregouet DA, et al. (November 2004). "Gender-specific association of a perilipin gene haplotype with obesity risk in a white population". Obesity Research. 12 (11): 1758–65. doi:10.1038/oby.2004.218. PMID 15601970.
- Corella D, Qi L, Sorlí JV, Godoy D, Portolés O, Coltell O, et al. (September 2005). "Obese subjects carrying the 11482G>A polymorphism at the perilipin locus are resistant to weight loss after dietary energy restriction". The Journal of Clinical Endocrinology and Metabolism. 90 (9): 5121–6. doi:10.1210/jc.2005-0576. PMID 15985482.
Further reading
- Brasaemle DL (December 2007). "Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis". Journal of Lipid Research. 48 (12): 2547–59. doi:10.1194/jlr.R700014-JLR200. PMID 17878492.
- Tai ES, Ordovas JM (April 2007). "The role of perilipin in human obesity and insulin resistance". Current Opinion in Lipidology. 18 (2): 152–6. doi:10.1097/MOL.0b013e328086aeab. PMID 17353663. S2CID 23086524.
- Nishiu J, Tanaka T, Nakamura Y (March 1998). "Isolation and chromosomal mapping of the human homolog of perilipin (PLIN), a rat adipose tissue-specific gene, by differential display method". Genomics. 48 (2): 254–7. doi:10.1006/geno.1997.5179. PMID 9521880.
- Souza SC, Muliro KV, Liscum L, Lien P, Yamamoto MT, Schaffer JE, et al. (March 2002). "Modulation of hormone-sensitive lipase and protein kinase A-mediated lipolysis by perilipin A in an adenoviral reconstituted system". The Journal of Biological Chemistry. 277 (10): 8267–72. doi:10.1074/jbc.M108329200. PMID 11751901.
- Hagström-Toft E, Qvisth V, Nennesmo I, Rydén M, Bolinder H, Enoksson S, et al. (December 2002). "Marked heterogeneity of human skeletal muscle lipolysis at rest". Diabetes. 51 (12): 3376–83. doi:10.2337/diabetes.51.12.3376. PMID 12453889.
- Mottagui-Tabar S, Rydén M, Löfgren P, Faulds G, Hoffstedt J, Brookes AJ, et al. (June 2003). "Evidence for an important role of perilipin in the regulation of human adipocyte lipolysis". Diabetologia. 46 (6): 789–97. doi:10.1007/s00125-003-1112-x. PMID 12802495.
- Wang Y, Sullivan S, Trujillo M, Lee MJ, Schneider SH, Brolin RE, et al. (August 2003). "Perilipin expression in human adipose tissues: effects of severe obesity, gender, and depot". Obesity Research. 11 (8): 930–6. doi:10.1038/oby.2003.128. PMID 12917496.
- Zhang HH, Souza SC, Muliro KV, Kraemer FB, Obin MS, Greenberg AS (December 2003). "Lipase-selective functional domains of perilipin A differentially regulate constitutive and protein kinase A-stimulated lipolysis". The Journal of Biological Chemistry. 278 (51): 51535–42. doi:10.1074/jbc.M309591200. PMID 14527948.
- Kern PA, Di Gregorio G, Lu T, Rassouli N, Ranganathan G (March 2004). "Perilipin expression in human adipose tissue is elevated with obesity". The Journal of Clinical Endocrinology and Metabolism. 89 (3): 1352–8. doi:10.1210/jc.2003-031388. PMID 15001633.
- Arvidsson E, Blomqvist L, Rydén M (May 2004). "Depot-specific differences in perilipin mRNA but not protein expression in obesity". Journal of Internal Medicine. 255 (5): 595–601. doi:10.1111/j.1365-2796.2004.01314.x. PMID 15078502. S2CID 10719527.
- Dalen KT, Schoonjans K, Ulven SM, Weedon-Fekjaer MS, Bentzen TG, Koutnikova H, et al. (May 2004). "Adipose tissue expression of the lipid droplet-associating proteins S3-12 and perilipin is controlled by peroxisome proliferator-activated receptor-gamma". Diabetes. 53 (5): 1243–52. doi:10.2337/diabetes.53.5.1243. PMID 15111493.
- Qi L, Corella D, Sorlí JV, Portolés O, Shen H, Coltell O, et al. (October 2004). "Genetic variation at the perilipin (PLIN) locus is associated with obesity-related phenotypes in White women". Clinical Genetics. 66 (4): 299–310. doi:10.1111/j.1399-0004.2004.00309.x. PMID 15355432. S2CID 24420287.
- Yan W, Chen S, Huang J, Shen Y, Qiang B, Gu D (November 2004). "Polymorphisms in PLIN and hypertension combined with obesity and lipid profiles in Han Chinese". Obesity Research. 12 (11): 1733–7. doi:10.1038/oby.2004.214. PMID 15601966.
- Qi L, Shen H, Larson I, Schaefer EJ, Greenberg AS, Tregouet DA, et al. (November 2004). "Gender-specific association of a perilipin gene haplotype with obesity risk in a white population". Obesity Research. 12 (11): 1758–65. doi:10.1038/oby.2004.218. PMID 15601970.
- Qi L, Tai ES, Tan CE, Shen H, Chew SK, Greenberg AS, et al. (June 2005). "Intragenic linkage disequilibrium structure of the human perilipin gene (PLIN) and haplotype association with increased obesity risk in a multiethnic Asian population". Journal of Molecular Medicine. 83 (6): 448–56. doi:10.1007/s00109-004-0630-4. PMID 15770500. S2CID 7820923.
- Forcheron F, Legedz L, Chinetti G, Feugier P, Letexier D, Bricca G, Beylot M (August 2005). "Genes of cholesterol metabolism in human atheroma: overexpression of perilipin and genes promoting cholesterol storage and repression of ABCA1 expression". Arteriosclerosis, Thrombosis, and Vascular Biology. 25 (8): 1711–7. CiteSeerX 10.1.1.581.2332. doi:10.1161/01.ATV.0000174123.19103.52. PMID 15961705. S2CID 5150107.
- Corella D, Qi L, Sorlí JV, Godoy D, Portolés O, Coltell O, et al. (September 2005). "Obese subjects carrying the 11482G>A polymorphism at the perilipin locus are resistant to weight loss after dietary energy restriction". The Journal of Clinical Endocrinology and Metabolism. 90 (9): 5121–6. doi:10.1210/jc.2005-0576. PMID 15985482.
- Moore HP, Silver RB, Mottillo EP, Bernlohr DA, Granneman JG (December 2005). "Perilipin targets a novel pool of lipid droplets for lipolytic attack by hormone-sensitive lipase". The Journal of Biological Chemistry. 280 (52): 43109–20. doi:10.1074/jbc.M506336200. PMID 16243839.
- Shimizu M, Akter MH, Emi Y, Sato R, Yamaguchi T, Hirose F, Osumi T (March 2006). "Peroxisome proliferator-activated receptor subtypes differentially cooperate with other transcription factors in selective transactivation of the perilipin/PEX11 alpha gene pair". Journal of Biochemistry. 139 (3): 563–73. doi:10.1093/jb/mvj053. PMID 16567422.