Pikaia
Pikaia gracilens is an extinct, primitive chordate animal known from the Middle Cambrian Burgess Shale of British Columbia. Described in 1911 by Charles Doolittle Walcott as an annelid, and in 1979 by Harry B. Whittington and Simon Conway Morris as a chordate, it became "one of the most famous early chordate fossils,"[1] or "famously known as the earliest described Cambrian chordate".[2] It is estimated to have lived during the latter period of the Cambrian explosion. Since its initial discovery, more than a hundred specimens have been recovered.[3]
| Pikaia | |
|---|---|
 | |
| Life reconstruction of Pikaia gracilens | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Family: | †Pikaiidae |
| Genus: | †Pikaia Walcott, 1911 |
| Species: | †P. gracilens |
| Binomial name | |
| †Pikaia gracilens Walcott, 1911 | |
The body structure resembles that of the lancelet and it swam perhaps much like an eel. A notochord and myomeres (segmented blocks of skeletal muscles) span the entire length of the body, and are considered the defining signatures of chordate characters. Its primitive nature is indicated by the body covering, a cuticle, which is characteristic of invertebrates and some protochordates.
The exact phylogenetic position is unclear. Proposed affinities include those of cephalochordata, craniata, or a stem-chordate not closely related to any extant lineage.[4] Popularly but falsely[5] attributed as an ancestor of all vertebrates,[6] or the oldest fish,[7] a close relative of the ancestor of humans,[8][9] it is generally viewed as a basal chordate alongside other Cambrian chordates; it is a close relative of vertebrate ancestors but it is not an ancestor itself.[2][10]
Discovery
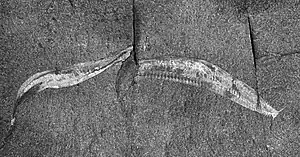
The fossils of Pikaia gracilens was discovered by Charles Walcott from the Burgess shale member of the Stephen formation in British Columbia, and described it in 1911. He named it after Pika Peak, a mountain in Alberta, Canada. Based on the obvious and regular segmentation of the body, as is the feature of annelids,[11] Walcott classified it as a polychaete worm and created a new family Pikaidae for it. (The family named was changed to Pikaiidae in 1962 by Princeton palaeontologist Benjamin Franklin Howell.[12]) Walcott was aware of the limitation of his classification, as he noted: "I am unable to place it within any of the families of the Polychaeta, owing to the absence of parapodia [paired protrusions on the sides of polychaete worms] on the body segments back of the fifth."[13]
University of Cambridge palaeontologist Harry B. Whittington and his student Simon Conway Morris re-examined the Burgess Shale fauna and noted the anatomical details of Pikaia for the first time. The fossil specimens bears features of notochord and muscle blocks that are fundamental structures of chordates, and not of annelids. Conway Morris presented a paper in 1977 that first indicated the possible chordate position but without further explanation.[14][15] He and Whittington were convinced that animal was obviously a chordate, as they wrote in the Scientific American in 1979:
Finally, we find among the Burgess Shale fauna one of the earliest-known invertebrate representatives of our own conspicuous corner of the animal kingdom: the chordate phylum... The chordates are represented in the Burgess Shale by the genus Pikaia and the single species P. gracilens.[16]
Conway Morris formally placed P. gracilens among the chordates in a paper in the Annual Review of Ecology and Systematics in 1979.[17] However, there were no structural analyses such as using microscopes to confirm the chordate features. The comparative description only earned a "putative" chordate status.[18] The chordate nature were often described with sceptical remarks for several decades.[19][20][21] It was only in 2012, when detailed analysis was reported by Conway Morris and Jean-Bernard Caron that the chordate position became generally accepted.[10][22]
The fossils are found only in a restricted series of horizons in the strata exposed on Fossil Ridge, close to the Yoho National Park. From the same location, other fish-like animal fossils named Metaspriggina were discovered in 1993. Conway Morris identified the animals as another Cambrian chordate.[23] The fossil specimens are preserved in the Smithsonian Institution and the Royal Ontario Museum.[3]
Description
Pikaia has a lancelet-like body, tapering at both ends, laterally flat and lacked a well-defined head. It measures an average of about 1+1⁄2 inches (3.8 cm) in length. Walcott recorded the longest individuals as 5 cm (2.0 in) in length.[13] Pikaia has a pair of large, antenna-like tentacles on its head that resembles those of invertebrates such as snails.[21] The attachment of the tentacles makes a two-lobed structure of the head.[24] The tentacles may be comparable to those in the present-day hagfish, a jawless chordate.[25] It has a small circular mouth that could be used to eat small food particles in a single bite. There are a series of short appendages on either side of the underside of the head just after the mouth, and their exact nature or function is unknown. The pharynx is associated with six pairs of slits with tiny filaments that could be used for respiratory apparatus.[26] In these ways, it differs from the modern lancelets, which have distinct pharyngeal gill slits on either sides of the pharynx and are used for filter feeding.[27]
A major primitive structure of Pikaia is a cuticle as its body covering. Cuticle is a hard protein layer predominantly found in invertebrates such as arthropods, molluscs, echinoderms and nematodes.[28][29] Unlike a typical cuticle, the cuticle of Pikaia does not have hard extracellular (exoskeleton) protection, and the entirely body is essentially soft-bodied.[30] Although primitive, Pikaia shows the essential prerequisites for vertebrates. When alive, Pikaia was a compressed, leaf-shaped animal with an expanded tail fin; the flattened body is divided into pairs of segmented muscle blocks, seen as faint vertical lines. The muscles lie on either side of a flexible structure resembling a rod that runs from the tip of the head to the tip of the tail.[31]
Pikaia was an active and free swimmer.[13] It likely swam by throwing its body into a series of S-shaped, zigzag curves, similar to the movement of eels; fish inherited the same swimming movement, but they generally have stiffer backbones. These adaptations may have allowed Pikaia to filter particles from the water as it swam along.[25] Pikaia was probably a slow swimmer, since it lacked the fast-twitch fibers that are associated with rapid swimming in modern chordates.[25]
Reinterpretations
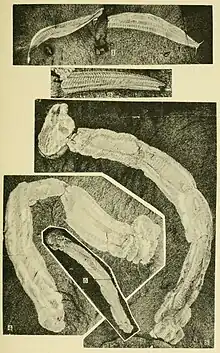
Walcott's original summary of the description of Pikaia reads:
Body elongate, slender, and tapering at each end. It is formed of many segments that are defined by strong annular shiny lines. Head small with two large eyes and two tentacles... Back of the head the first five segments carry short parapodia that appear to be divided into two parts. The enteric canal extends from end to end without change in character... This was one of the active, free-swimming annelids that suggest the Nephthydidae of the Polychaeta.[13]
Whittington and Conway Morris were the first to realise that Walcott's description and classification were not reliable and mostly inaccurate. They compared the body segments as described by Walcott with living animals and found that they were similar to the muscle bundles of chordates such as the living Amphioxus (Branchiostoma) as well as fishes, and not to superficial segments of annelids. They pictured that the muscles would be essential for swimming in water in wriggling motions. The enteric canal as observed by Walcott was not an ordinary digestive tract, it runs along with a stiff rod that resembles a notochord. They reported in 1979: "Although Pikaia differs from Amphioxus, in several important respects, the conclusion is that it is not a worm but a chordate appears inescapable."[16]
Conway Morris was convinced that the longitudinal rod was a notochord and the segments were muscle blocks that he concluded that Pikaia "is a primitive chordate rather than a polychaete. The earliest fish scales are Upper Cambrian, and Pikaia may not be far removed from the ancestral fish."[17] In 1982, he added further description in his Atlas of the Burgess Shale that Pikaia had one or more fins, but did not specify where they were present.[10]
Pikaia was not popularly known as a chordate fossil or as an ancient chordate until 1989.[32] That year, Harvard University palaeontologist Stephen Jay Gould wrote in his book Wonderful Life: The Burgess Shale and the Nature of History: "Pikaia is not an annelid worm. It is a chordate, a member of our own phylum—in fact, the first recorded member of our immediate ancestry."[33] From this remark Pikaia became generally recognised as a chordate and ancestor of vertebrates.[22][34][35]
In 1993, Conway Morris came up with another possible chordate feature. He identified structures that looked like gill slits but gave a cautious remark: "[They] may have been present, but are hard to identify with certainty in the compressed material available.[24] The tiny pores on the side of the pharynx are normally gill slits in living chordates.[36] He also noticed that Pikaia is similar to Amphioxus in most general aspects, with major difference in its notochord not reaching the anterior end.[24]
Not all palaeontologists were convinced of the chordate designation without better analysis.[19][20] In 2001, Nicholas D. Holland from the Scripps Institution of Oceanography and Junyuan Chen from the Chinese Academy of Sciences criticised the presentation in Wonderful Life, saying that the "reinterpretation [of Pikaia as a chordate] became almost universally accepted after its unqualified and forceful endorsement by Gould"; concluding that "the cephalochordate affinity of Pikaia is at best only weakly indicated by the characters visible in fossils discovered so far."[37] In 2010, an international team of palaeontologists argued that Pikaia has sufficiently invertebrate characters, and that it mostly look like a much younger extinct animal, the Tully monster (Tullimonstrum gregarium),[21] which is still debated as either an invertebrate or a chordate.[38][39]
Another component of Pikaia fossils that constrains the animal to be accepted as a chordate is its distinct invertebrate character; its preservational mode suggests that it had cuticle. The cuticle as a body covering is uncharacteristic of the vertebrates,[40] but is a dominant feature of invertebrates.[28] The presence of earlier chordates among the Chengjiang, including Haikouichthys and Myllokunmingia, appears to show that cuticle is not necessary for preservation, overruling the taphonomic argument,[41] but the presence of tentacles remains intriguing, and the organism cannot be assigned conclusively, even to the vertebrate stem group. Its anatomy closely resembles the modern creature Branchiostoma.[42]
A fossil species Myoscolex ateles, discovered in 1979 from Cambrian Emu Bay shale of Kangaroo Island in South Australia, had been debated as among the oldest annelids, or at least other invertebrate groups.[43][44] Polish palaeontologist Jerzy Dzik in his formal description in 2003 notes that it "closely resembles the slightly geologically younger Pikaia" in having smooth cuticle as well as muscular segmentation, and projections on its backside (ventral chaetae) that look like Pikaia's tentacles. He concluded:
In fact, there is little evidence for chordate affinities of Pikaia. Its relationship with Myoscolex [as annelid in his proposition] appears a much better solution. Both were initially identified as polychaetes and this line of inference perhaps deserves confrontation with more recent evidence than that available to the authors who proposed these genera.[45]
Comprehensive description
.svg.png.webp)
The first comprehensive description of Pikaia was published by Conway Morris and Jean-Bernard Caron in the May 2012 issue of Biological Reviews. The anatomical examination and interpretation based on 114 fossil specimens confirm the classification as a chordate.[3] According to the new assessment, Pikaia fossils indicate important features that define the animal as a primitive chordate.[10] All Pikaia fossils are in the range of 1.5 to 6 cm in length, with an average of 4 cm. Having a laterally compressed and fusiform (tapering at both ends) body, the exact width and height are variable, and normally its height is twice that of its width throughout it body.[3]
The head is bilaterally symmetrical with a distinct pair of tentacles. Due to its small size, only about 1 mm in diameter, the structural details are indistinguishable. Some specimens show a darker central line on the tentacles which may represent a nervous fibre; thus making the tentacles as sensory feelers.[46] A mouth is marked by a small opening at the anterior end of the gut towards the underside of the head. There are no jaws and teeth. Walcott had mentioned the presence of two large eyes,[13] but no specimens, including Walcott's original collection, show any evidence of eyes.[3]
One of the most unusual body parts is a series of appendages just posterior to the tentacles. Walcott had called the appendages parapodia, as a kind of body protrusions that aid locomotion in snails,[47] and mentioned five parapodia in each individual.[13] He was even puzzled by the absence on the major part of the body.[13] Other specimens have up to nine such appendages, and could not be parapodia. Fins are present as an expansion of the body on the dorsal and ventral sides. They are not present in many specimens indicating that they are delicate membranes and were lost during fossilisation.[3]
The backside of Pikaia fossils show a hollow tubular structure that extends throughout most of the body length, but not the anterior region. It is easily noticeable as a highly light-reflective portion and is known as the dorsal organ. Once described as the notochord,[16][17] its nature is not yet fully resolved and could be a storage organ. The true notochord, along with a nerve chord, is a fine lateral line that runs just beneath the thick dorsal organ.[3]
The main chordate character is a series of myomeres that extends from the anterior to the posterior region. On average, there are 100 such myomeres in each individual. The muscle segments are not simply "annular shiny lines" as Walcott described,[13] but are in concentric bends in the form of V-shaped chevron. The myomeres at the anterior end as simpler in appearance and show circular arrangement. Conwy Morris and Caron concluded:
Whilst the possibility that Pikaia is simply convergent on the chordates cannot be dismissed, we prefer to build a scenario that regards Pikaia as the most stem-ward of the chordates with links to the phylogenetically controversial yunnanozoans. This hypothesis has implications for the evolution of the myomeres, notochord and gills.[3]
Evolutionary importance

Much debate on whether Pikaia is a vertebrate ancestor, its worm-like appearance notwithstanding, exists in scientific circles. It looks like a worm that has been flattened sideways (lateral compression). The fossils compressed within the Burgess Shale show chordate features such as traces of an elongate notochord, dorsal nerve cord, and blocks of muscles (myotomes) down either side of the body – all critical features for the evolution of the vertebrates.
The notochord, a flexible rod-like structure that runs along the back of the animal, lengthens and stiffens the body so that it can be flexed from side to side by the muscle blocks for swimming. In the fish and all subsequent vertebrates, the notochord forms the backbone (or vertebral column). The backbone strengthens the body, supports strut-like limbs, and protects the vital dorsal nerve cord, while at the same time allowing the body to bend.
A Pikaia lookalike, the lancelet Branchiostoma, still exists today. With a notochord and paired muscle blocks, the lancelet and Pikaia belong to the chordate group of animals from which the vertebrates descended. Molecular studies have refuted earlier hypotheses that lancelets might be the closest living relative to the vertebrates, instead favoring tunicates in this position;[48] other extant and fossil groups, such as acorn worms and graptolites, are more primitive.[lower-alpha 1]
The presence of cuticle, one of the principal characters of higher invertebrates,[28][29] in Pikaia can be understood from the evolutionary trends. A Cambrian invertebrate, Myoscolex ateles was described to be structurally similar to Pikaia particularly in having smooth cuticle as well as muscular segmentation, and projections on its backside (ventral chaetae) that look like Pikaia's tentacles.[45] Although chordates normally lack the cuticle, a type of cuticle is present in some cephalochordates, indicating that primitive characters are retained in lower chordates.[49]
Subsequently, Mallatt and Holland reconsidered Conway Morris and Caron's description, and concluded that many of the newly recognized characters are unique, already-divergent specializations that would not be helpful for establishing Pikaia as a basal chordate.[10]
Development of the head
The first sign of head development, cephalization, is seen in chordates such as Pikaia and Branchiostoma. It is thought that development of a head structure resulted from a long body shape, a swimming habit, and a mouth at the end that came into contact with the environment first, as the animal swam forward. The search for food required ways of continually testing what lay ahead so it is thought that anatomical structures for seeing, feeling, and smelling developed around the mouth. The information these structures gathered was processed by a swelling of the nerve cord (efflorescence) – the precursor of the brain. Altogether, these front-end structures formed the distinct part of the vertebrate body known as the head.[31]
Evolutionary interpretation
Once thought to be closely related to the ancestor of all vertebrates, Pikaia has received particular attention among the multitude of animal fossils found in the famous Burgess Shale and other Cambrian fauna. In 1979, Whittington and Conway Morris first explained the evolutionary importance of Pikaia. Realising the fossil to be that of a chordate in the Cambrian rocks, chordates could have originated much earlier than expected, as they commented: "The superb preservation of this Middle Cambrian organism [Pikaia] makes it a landmark history of the phylum [Chordata] to which all vertebrates, including man, belong."[16] It is for this knowledge Pikaia as an old chordate that it is often misleadingly and falsely[5] attributed to as an ancestor of all vertebrates,[6] or the oldest fish,[7] or the oldest ancestor of humans.[8][9]
Before Pikaia and other Cambrian chordates were fully appreciated, it was generally believed that the first chordates appeared much later, such as in Ordovician (484–443 mya). The establishment of Cambrian chordates, according to Stephen Jay Gould, prompted "revised views of evolution, ecology and development," and remarked: "So much for chordate uniqueness marked by slightly later evolution."[50] However, Gould did not believe that Pikaia itself was unique as an early chordate or that it was "the actual ancestor of vertebrates;" he presumed that there could be undiscovered fossils that are more closely linked to vertebrate ancestry.[51]
The presence of a creature as complex as Pikaia some 530 million years ago reinforces the controversial view that the diversification of life must have extended back well before Cambrian times – perhaps deep into the Precambrian.[31] The discovery of Metaspriggina, a primitive fish of similar age which seems to have already started to develop a jaw and the presence of conodonts, the teeth of an extinct type of fish belonging to the Agnatha some 20 Ma before Pikaia, does provide support for that view.
Gould's interpretation and evolutionary contingency
Gould, in his presidential address of the Paleontological Society on 27 October 1988, cited Pikaia to explain the trends of evolutionary changes:
Wind back life's tape to the Burgess (first erasing what actually came after), let it play again, and this time a quite different cast may emerge. If the cast lacked Pikaia, the first chordate, we might not be here—and the world would be no worse... Let us thank our lucky stars for the survival of Pikaia.[52]
He elaborated the same idea in "An epilogue on Pikaia" in his book Wonderful Life "to save the best for the last," in which he made a statement:
Pikaia is the missing and final link in our story of contingency—the direct connection between Burgess decimation and eventual human evolution... Wind the tape of life back to Burgess times, and let it play again. If Pikaia does not survive in the replay, we are wiped out of future history—all of us, from shark to robin to orangutan... And so, if you wish to ask the question of the age—why do humans exist?—a major part of the answer, touching those aspects of the issue that science can treat at all, must be: because Pikaia survived the Burgess decimation.[51]
This interpretation that the chances of evolutionary products are unpredictable is known as evolutionary contingency.[53][54] Gould, from this statement, is regarded as "the most famous proponent" of the concept.[55] His idea has inspired many research involving enolutionary contingency from palaeontology to molecular biology.[56][57][58] He used Pikaia among the Cambrian animals as an epitome of contingent event in the entire evolution of life; if Pikaia had not existed, the rest of chordate animals might not have evolved, thus completely changing the diversity of life as we know. According to him, contingency is a major factor that drives large-scale evolution (macroevolution) and dictates that evolution has no inevitable destiny or outcome.[59] However, as Gould explained, "The bad news is that we can't possibly perform the experiment."[58]
Notes
- Acorn worms are classified as hemichordates, which have a notochord-like structure only at the early stage of their lives.
References
- Gee, Henry (2018). Across the Bridge: Understanding the Origin of the Vertebrates. University of Chicago Press. p. 83. ISBN 978-0-226-40319-9.
- McMenamin, Mark A. S. (2019). "Cambrian Chordates and Vetulicolians". Geosciences. 9 (8): 354. Bibcode:2019Geosc...9..354M. doi:10.3390/geosciences9080354. ISSN 2076-3263.
- Morris, Simon Conway; Caron, Jean-Bernard (2012). "Pikaia gracilens Walcott, a stem-group chordate from the Middle Cambrian of British Columbia". Biological Reviews. 87 (2): 480–512. doi:10.1111/j.1469-185X.2012.00220.x. PMID 22385518. S2CID 27671780.
- "Pikaia gracilens" Burgess Shale Fossil Gallery. Virtual Museum of Canada. 2011.
- Danchin, Etienne G.J.; Pontarotti, Pierre (2004). "Towards the reconstruction of the bilaterian ancestral pre-MHC region". Trends in Genetics. 20 (12): 587–591. doi:10.1016/j.tig.2004.09.009. PMID 15522451.
- SciTechDaily (2012-03-06). "Pikaia Gracilens Is a Relative of but not the Genetic Ancestor to All Vertebrates". SciTechDaily. Retrieved 2022-09-23.
- "An Early Chordate that Lived 513 Million Years Ago". Discover Magazine. Retrieved 2022-09-23.
- "Worm-like creature could be a relative of the earliest vertebrate". 2012-03-05. Retrieved 2022-09-23.
- "Early Vertibrate Relative found". University of Cambridge. 2012-03-06. Retrieved 2022-09-23.
- Mallatt, Jon; Holland, Nicholas (2013). "Pikaia gracilens Walcott: Stem Chordate, or Already Specialized in the Cambrian?". Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 320 (4): 247–271. doi:10.1002/jez.b.22500. PMID 23606659.
- Balavoine, Guillaume (2014). "Segment formation in Annelids: patterns, processes and evolution". The International Journal of Developmental Biology. 58 (6–8): 469–483. doi:10.1387/ijdb.140148gb. ISSN 1696-3547. PMID 25690963.
- Howell, B. F. (1962). "Worms". In Moore (ed.). Treatise on Invertebrate Paleontology Part W. Miscellanea (R. C. ed.). New York: Geological Society of America and University of Kansas Press. pp. W144–W177. ISBN 978-0-8137-3024-0.
- Walcott, Charles D. (1911). "Cambrian Geology and Paleontology II: No.5--Middle Cambrian Annelids" (PDF). Smithsonian Miscellaneous Collections. 57 (5): 109–144.
- Simon, Conway Morris (1977). "Aspects of the Burgess Shale fauna, with particular reference to the non-arthropod component". Journal of Paleontology. 51 (Suppl 2): 7–8.
- Whittington, H.B. (1980). "The significance of the fauna of the Burgess Shale, Middle Cambrian, British Columbia". Proceedings of the Geologists' Association. 91 (3): 127–148. Bibcode:1980PrGA...91..127W. doi:10.1016/S0016-7878(80)80034-4.
- Morris, Simon Conway; Whittington, H. B. (1979). "The Animals of the Burgess Shale". Scientific American. 241 (1): 122–135. Bibcode:1979SciAm.241a.122M. doi:10.1038/scientificamerican0779-122. ISSN 0036-8733. JSTOR 24965247.
- Morris, Simon Conway (1979). "The Burgess Shale (Middle Cambrian) Fauna". Annual Review of Ecology and Systematics. 10: 327–349. doi:10.1146/annurev.es.10.110179.001551. ISSN 0066-4162. JSTOR 2096795.
- Butterfield, Nicholas J. (1990). "Organic preservation of non-mineralizing organisms and the taphonomy of the Burgess Shale". Paleobiology. 16 (3): 272–286. Bibcode:1990Pbio...16..272B. doi:10.1017/S0094837300009994. ISSN 0094-8373. S2CID 133486523.
- Insom, Emilio; Pucci, Antonella; Simonetta, Alberto M. (1995). "Cambrian Protochordata, their origin and significance". Bolletino di Zoologia. 62 (3): 243–252. doi:10.1080/11250009509356072. ISSN 0373-4137.
- Hall, Brian K. (1992), Hall, Brian K. (ed.), "The Burgess Shale: Different types of animals", Evolutionary Developmental Biology, Dordrecht: Springer Netherlands, pp. 13–37, doi:10.1007/978-94-015-7926-1_2, ISBN 978-94-015-7926-1, retrieved 2022-09-23
- Turner, Susan; Burrow, Carole J.; Schultze, Hans-Peter; Blieck, Alain; Reif, Wolf-Ernst; Rexroad, Carl B.; Bultynck, Pierre; Nowlan, Godfrey S. (2010). "False teeth: conodont-vertebrate phylogenetic relationships revisited". Geodiversitas. 32 (4): 545–594. doi:10.5252/g2010n4a1. ISSN 1280-9659. S2CID 86599352.
- Janvier, Philippe (2015). "Facts and fancies about early fossil chordates and vertebrates". Nature. 520 (7548): 483–489. Bibcode:2015Natur.520..483J. doi:10.1038/nature14437. ISSN 1476-4687. PMID 25903630. S2CID 4462904.
- Conway Morris, Simon (2008). "A Redescription of a Rare Chordate, Metaspriggina walcotti Simonetta and Insom, from the Burgess Shale (Middle Cambrian), British Columbia, Canada". Journal of Paleontology. 82 (2): 424–430. Bibcode:2008JPal...82..424M. doi:10.1666/06-130.1. ISSN 0022-3360. S2CID 85619898.
- Morris, S. Conway (1993). "The fossil record and the early evolution of the Metazoa". Nature. 361 (6409): 219–225. Bibcode:1993Natur.361..219M. doi:10.1038/361219a0. ISSN 0028-0836. S2CID 86276440.
- Lacalli, T. (2012). "The Middle Cambrian fossil Pikaia and the evolution of chordate swimming". EvoDevo. 3 (1): 12. doi:10.1186/2041-9139-3-12. PMC 3390900. PMID 22695332.
- Striedter, Georg F.; Northcutt, R. Glenn (2020). Brains Through Time: A Natural History of Vertebrates. Oxford University Press. p. 70. ISBN 978-0-19-512568-9.
- Wang, Wenjun; Wang, Changliu; Chen, Wei; Ding, Shuo (2021). "Advances in immunological research of amphioxus". Developmental & Comparative Immunology. 118: 103992. doi:10.1016/j.dci.2020.103992. PMID 33387559. S2CID 230282227.
- Wilt, Fred H.; Killian, Christopher E.; Livingston, Brian T. (2003). "Development of calcareous skeletal elements in invertebrates". Differentiation. 71 (4–5): 237–250. doi:10.1046/j.1432-0436.2003.7104501.x. PMID 12823225.
- Giribet, G.; Ribera, C. (1998). "The position of arthropods in the animal kingdom: a search for a reliable outgroup for internal arthropod phylogeny". Molecular Phylogenetics and Evolution. 9 (3): 481–488. doi:10.1006/mpev.1998.0494. ISSN 1055-7903. PMID 9667996.
- Parry, Luke A.; Smithwick, Fiann; Nordén, Klara K.; Saitta, Evan T.; Lozano-Fernandez, Jesus; Tanner, Alastair R.; Caron, Jean-Bernard; Edgecombe, Gregory D.; Briggs, Derek E. G.; Vinther, Jakob (2018). "Soft-Bodied Fossils Are Not Simply Rotten Carcasses - Toward a Holistic Understanding of Exceptional Fossil Preservation: Exceptional Fossil Preservation Is Complex and Involves the Interplay of Numerous Biological and Geological Processes". BioEssays. 40 (1): 1700167. doi:10.1002/bies.201700167. PMID 29193177. S2CID 39128197.
- Palmer, D., (2000). The Atlas of the Prehistoric World. London: Marshall Publishing Ltd. p66-67.
- Powell, Russell (2020-02-04). Contingency and Convergence: Toward a Cosmic Biology of Body and Mind. MIT Press. p. 62. ISBN 978-0-262-35660-2.
- Gould, Stephen Jay (2000) [1989]. Wonderful Life: The Burgess Shale and the Nature of History. Vintage/W.W. Norton & Company. p. 321. ISBN 978-0-09-927345-5. OCLC 18983518.
- Beatty, John (2016). "What are narratives good for?". Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences. 58: 33–40. doi:10.1016/j.shpsc.2015.12.016. PMID 26806602.
- Schubert, Michael; Escriva, Hector; Xavier-Neto, José; Laudet, Vincent (2006). "Amphioxus and tunicates as evolutionary model systems". Trends in Ecology & Evolution. 21 (5): 269–277. doi:10.1016/j.tree.2006.01.009. PMID 16697913.
- Ruppert, Edward E (2005). "Key characters uniting hemichordates and chordates: homologies or homoplasies?". Canadian Journal of Zoology. 83 (1): 8–23. doi:10.1139/z04-158. ISSN 0008-4301.
- Holland, N. D.; Chen, J. (2001). "Origin and early evolution of the vertebrates: new insights from advances in molecular biology, anatomy, and palaeontology". BioEssays. 23 (2): 142–151. doi:10.1002/1521-1878(200102)23:2<142::AID-BIES1021>3.0.CO;2-5. ISSN 0265-9247. PMID 11169587. S2CID 25572329.
- Sallan, Lauren; Giles, Sam; Sansom, Robert S.; Clarke, John T.; Johanson, Zerina; Sansom, Ivan J.; Janvier, Philippe (2017). Zhang, Xi-Guang (ed.). "The 'Tully Monster' is not a vertebrate: characters, convergence and taphonomy in Palaeozoic problematic animals". Palaeontology. 60 (2): 149–157. Bibcode:2017Palgy..60..149S. doi:10.1111/pala.12282. S2CID 90132820.
- McCoy, Victoria E.; Wiemann, Jasmina; Lamsdell, James C.; Whalen, Christopher D.; Lidgard, Scott; Mayer, Paul; Petermann, Holger; Briggs, Derek E. G. (2020). "Chemical signatures of soft tissues distinguish between vertebrates and invertebrates from the Carboniferous Mazon Creek Lagerstätte of Illinois". Geobiology. 18 (5): 560–565. Bibcode:2020Gbio...18..560M. doi:10.1111/gbi.12397. ISSN 1472-4669. PMID 32347003. S2CID 216646333.
- Butterfield, N. J. (1990), "Organic preservation of non-mineralizing organisms and the taphonomy of the Burgess Shale", Paleobiology, 16 (3): 272–286, Bibcode:1990Pbio...16..272B, doi:10.1017/S0094837300009994, JSTOR 2400788, S2CID 133486523
- Conway Morris, S. (2008), "A Redescription of a Rare Chordate, Metaspriggina walcotti Simonetta and Insom, from the Burgess Shale (Middle Cambrian), British Columbia, Canada", Journal of Paleontology, 82 (2): 424–430, Bibcode:2008JPal...82..424M, doi:10.1666/06-130.1, S2CID 85619898, retrieved 2009-04-28
- Donoghue, P. C. J.; Purnell, M. A. (2005), "Genome duplication, extinction and vertebrate evolution" (PDF), Trends in Ecology & Evolution, 20 (6): 312–319, doi:10.1016/j.tree.2005.04.008, PMID 16701387, archived from the original (PDF) on 2008-12-17, retrieved 2008-11-06
- Briggs, Derek E. G.; Nedin, Christopher (1997). "The taphonomy and affinities of the problematic fossil Myoscolex from the Lower Cambrian Emu Bay Shale of South Australia". Journal of Paleontology. 71 (1): 22–32. Bibcode:1997JPal...71...22B. doi:10.1017/S0022336000038919. ISSN 0022-3360. S2CID 131851540.
- Morris, Simon Conway; Peel, John S. (2008). "The Earliest Annelids: Lower Cambrian Polychaetes from the Sirius Passet Lagerstätte, Peary Land, North Greenland". Acta Palaeontologica Polonica. 53 (1): 137–148. doi:10.4202/app.2008.0110. ISSN 0567-7920. S2CID 35811524.
- Dzik, Jerzy (2004). "Anatomy and relationships of the Early Cambrian worm Myoscolex". Zoologica Scripta. 33 (1): 57–69. doi:10.1111/j.1463-6409.2004.00136.x. ISSN 0300-3256. S2CID 85216629.
- Hsieh, Shannon; Plotnick, Roy E.; Bush, Andrew M. (2022). "The Phanerozoic aftermath of the Cambrian information revolution: sensory and cognitive complexity in marine faunas". Paleobiology. 48 (3): 397–419. Bibcode:2022Pbio...48..397H. doi:10.1017/pab.2021.46. ISSN 0094-8373. S2CID 246399509.
- Özpolat, B. Duygu; Bely, Alexandra E. (2016). "Developmental and molecular biology of annelid regeneration: a comparative review of recent studies". Current Opinion in Genetics & Development. 40: 144–153. doi:10.1016/j.gde.2016.07.010. ISSN 1879-0380. PMID 27505269.
- Delsuc; et al. (2008). "Additional molecular support for the new chordate phylogeny" (PDF). Genesis. 46 (11): 592–604. doi:10.1002/dvg.20450. PMID 19003928. S2CID 205771088.
- Igawa, Takeshi; Nozawa, Masafumi; Suzuki, Daichi G.; Reimer, James D.; Morov, Arseniy R.; Wang, Yiquan; Henmi, Yasuhisa; Yasui, Kinya (2017-04-25). "Evolutionary history of the extant amphioxus lineage with shallow-branching diversification". Scientific Reports. 7 (1): 1157. Bibcode:2017NatSR...7.1157I. doi:10.1038/s41598-017-00786-5. ISSN 2045-2322. PMC 5430900. PMID 28442709.
- Gould, Stephen Jay (1995). "Of it, not above it". Nature. 377 (6551): 681–682. Bibcode:1995Natur.377..681G. doi:10.1038/377681a0. ISSN 0028-0836. S2CID 5589793.
- Gould, Stephen Jay (1989). Ibid. pp. 322–323. OCLC 45316756.
- Gould, Stephen Jay (1988). "Trends as changes in variance: a new slant on progress and directionality in evolution". Journal of Paleontology. 62 (3): 319–329. Bibcode:1988JPal...62..319G. doi:10.1017/S0022336000059126. JSTOR 1305411. S2CID 222252369.
- Blaser, Kent (1999). "The History of Nature and the Nature of History: Stephen Jay Gould on Science, Philosophy, and History". The History Teacher. 32 (3): 411–430. doi:10.2307/494379. ISSN 0018-2745. JSTOR 494379.
- Blount, Zachary D. (2016). "A case study in evolutionary contingency". Studies in History and Philosophy of Biological and Biomedical Sciences. 58: 82–92. doi:10.1016/j.shpsc.2015.12.007. ISSN 1879-2499. PMID 26787098.
- Erwin, Douglas H. (2006). "Evolutionary contingency". Current Biology. 16 (19): R825–826. doi:10.1016/j.cub.2006.08.076. ISSN 0960-9822. PMID 17027471. S2CID 5649275.
- Harms, Michael J.; Thornton, Joseph W. (2014). "Historical contingency and its biophysical basis in glucocorticoid receptor evolution". Nature. 512 (7513): 203–207. Bibcode:2014Natur.512..203H. doi:10.1038/nature13410. ISSN 0028-0836. PMC 4447330. PMID 24930765.
- Xie, Victoria Cochran; Pu, Jinyue; Metzger, Brian Ph; Thornton, Joseph W.; Dickinson, Bryan C. (2021). "Contingency and chance erase necessity in the experimental evolution of ancestral proteins". eLife. 10: e67336. doi:10.7554/eLife.67336. ISSN 2050-084X. PMC 8282340. PMID 34061027.
- Blount, Zachary D.; Lenski, Richard E.; Losos, Jonathan B. (2018). "Contingency and determinism in evolution: Replaying life's tape". Science. 362 (6415): eaam5979. Bibcode:2018Sci...362M5979B. doi:10.1126/science.aam5979. ISSN 1095-9203. PMID 30409860. S2CID 53239705.
- Briggs, Derek E. G. (2002). "Stephen Jay Gould (1941–2002)". Nature. 417 (6890): 706. doi:10.1038/417706a. ISSN 1476-4687. S2CID 4356596.
Further reading
- Bishop, A., Woolley, A. and Hamilton, W. (1999) Minerals, Rocks and Fossils. London: Phillip's
- Conway Morris, Simon. 1998. The Crucible of Creation: The Burgess Shale and the Rise of Animals. Oxford University Press, New York, New York.
- Norman, D. (1994) Prehistoric Life: the Rise of the Vertebrates, London: Boxtree
- Sheldon, P., Palmer D., Spicer, B. (2001). Fossils and the History of Life. Aberystwyth: Cambrian Printers/The Open University. p. 41-42.
External links
- "Pikaia gracilens". Burgess Shale Fossil Gallery. Virtual Museum of Canada. 2011. Archived from the original on 2020-11-12.
- La evolución de las especies: ¿por qué sobrevivió Pikaia? (Spanish)
- Fossils of the Burgess Shale - Middle Cambrian
- Pikaia gracilens Walcott, a stem-group chordate from the Middle Cambrian of British Columbia
| Forerunners |
|
| |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jawless fish |
| ||||||||||||
| Jawed fish |
| ||||||||||||
| Lists | |||||||||||||
| Related | |||||||||||||
† extinct | |||||||||||||
