Wood–Ljungdahl pathway
The Wood–Ljungdahl pathway is a set of biochemical reactions used by some bacteria. It is also known as the reductive acetyl-coenzyme A (Acetyl-CoA) pathway.[1] This pathway enables these organisms to use hydrogen as an electron donor, and carbon dioxide as an electron acceptor and as a building block for biosynthesis.
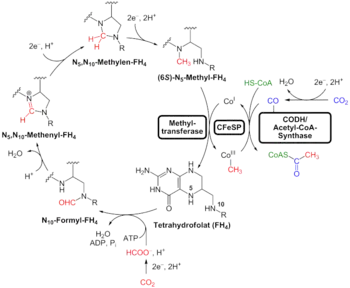
In this pathway carbon dioxide is reduced to carbon monoxide and formic acid or directly into a formyl group, the formyl group is reduced to a methyl group and then combined with the carbon monoxide and Coenzyme A to produce acetyl-CoA. Two specific enzymes participate on the carbon monoxide side of the pathway: CO Dehydrogenase and acetyl-CoA synthase. The former catalyzes the reduction of the CO2 and the latter combines the resulting CO with a methyl group to give acetyl-CoA.[1][2]
Some anaerobic bacteria use the Wood–Ljungdahl pathway in reverse to break down acetate. For example, Sulfate reducing bacteria oxidize acetate completely to CO2 and H2 coupled with the reduction of sulfate to sulfide.[3] When operating in the reverse direction, the acetyl-CoA synthase is sometimes called acetyl-CoA decarbonylase.
Not to be confused with the Wood-Ljungdahl pathway, an evolutionarily related but biochemically distinct pathway named the Wolfe Cycle[4] occurs exclusively in some methanogenic archaea called methanogens.[5] In these anaerobic archaea, the Wolfe Cycle functions as a methanogenesis pathway to reduce CO2 into methane with electron donors such as hydrogen and formate.[6] Similar to the reverse Krebs cycle and the Calvin cycle, the Wolfe Cycle is cyclic, but the Wood-Ljungdahl pathway is not.
Evolution
Relevance to abiogenesis
It has been proposed that the reductive acetyl-CoA pathway might have begun at deep sea alkaline hydrothermal vents where metal sulfides and metals catalyze the prebiotic reactions of the reductive acetyl-CoA pathway.[7] Recent experiments have tried to replicate this pathway by attempting to reduce CO2, with very little pyruvate observed using native iron as a reducing agent (<0.03 mM),[8] and even less so under hydrothermal settings with H2 (10 μM).[9] Joseph Moran and colleagues state that "it has been proposed that either the complete or “horseshoe” forms of the rTCA cycle may have once been united with the acetyl CoA pathway in an ancestral, possibly prebiotic, carbon fixation network".[8]
Last universal common ancestor
A 2016 study of the genomes of a set of bacteria and archaea suggested that the last universal common ancestor (LUCA) of all cells was using an ancient Wood–Ljungdahl pathway in a hydrothermal setting,[10] but more recent work challenges this conclusion as they argued that the previous study had "undersampled protein families, resulting in incomplete phylogenetic trees which do not reflect protein family evolution".[11] However geological evidence and phylogenomic reconstructions of the metabolic network of the common ancestors of archaea and bacteria support that LUCA fixed CO2 and relied on H2.[12][9]
References
- Ragsdale Stephen W (2006). "Metals and Their Scaffolds To Promote Difficult Enzymatic Reactions". Chem. Rev. 106 (8): 3317–3337. doi:10.1021/cr0503153. PMID 16895330.
- Paul A. Lindahl "Nickel-Carbon Bonds in Acetyl-Coenzyme A Synthases/Carbon Monoxide Dehydrogenases" Met. Ions Life Sci. 2009, volume 6, pp. 133–150. doi:10.1039/9781847559159-00133
- Spormann, Alfred M.; Thauer, Rudolf K. (1988). "Anaerobic acetate oxidation to CO2 by Desulfotomaculum acetoxidans". Archives of Microbiology. 150 (4): 374–380. doi:10.1007/BF00408310. ISSN 0302-8933. S2CID 2158253.
- Thauer, Rudolf K. (2012). "The Wolfe cycle comes full circle". Proceedings of the National Academy of Sciences of the United States of America. 109 (38): 15084–15085. doi:10.1073/pnas.1213193109. PMC 3458314. PMID 22955879.
- Matschiavelli, N.; Oelgeschlager, E.; Cocchiararo, B.; Finke, J.; Rother, M. (2012). "Function and regulation of isoforms of carbon monoxide dehydrogenase/acetyl-CoA synthase in Methanosarcina acetivorans". Journal of Bacteriology. 194 (19): 5377–87. doi:10.1128/JB.00881-12. PMC 3457241. PMID 22865842.
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, WB. (2018). "Methanogenesis". Current Biology. 28 (13): R727–R732. doi:10.1016/j.cub.2018.05.021. PMID 29990451.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Russell, M. J.; Martin, W. (2004). "The rocky roots of the acetyl-CoA pathway". Trends in Biochemical Sciences. 29 (7): 358–363. doi:10.1016/j.tibs.2004.05.007. ISSN 0968-0004. PMID 15236743.
- Varma, Sreejith J.; Muchowska, Kamila B.; Chatelain, Paul; Moran, Joseph (2018-04-23). "Native iron reduces CO2 to intermediates and end-products of the acetyl-CoA pathway". Nature Ecology & Evolution. 2 (6): 1019–1024. doi:10.1038/s41559-018-0542-2. ISSN 2397-334X. PMC 5969571. PMID 29686234.
- Preiner, Martina; Igarashi, Kensuke; Muchowska, Kamila B.; Yu, Mingquan; Varma, Sreejith J.; Kleinermanns, Karl; Nobu, Masaru K.; Kamagata, Yoichi; Tüysüz, Harun; Moran, Joseph; Martin, William F. (April 2020). "A hydrogen-dependent geochemical analogue of primordial carbon and energy metabolism". Nature Ecology & Evolution. 4 (4): 534–542. doi:10.1038/s41559-020-1125-6. ISSN 2397-334X. PMID 32123322. S2CID 211729738.
- M. C. Weiss; et al. (2016). "The physiology and habitat of the last universal common ancestor". Nature Microbiology. 1 (16116): 16116. doi:10.1038/nmicrobiol.2016.116. PMID 27562259. S2CID 2997255.
- S. J. Berkemer; et al. (2021). "A new analysis of archaea-bacteria domain separation: Variable phylogenetic distance and the tempo of early evolution". Molecular Biology and Evolution. 37 (8): 2332–2340. doi:10.1093/molbev/msaa089. PMC 7403611. PMID 32316034.
- Xavier, Joana C.; Gerhards, Rebecca E.; Wimmer, Jessica L. E.; Brueckner, Julia; Tria, Fernando D. K.; Martin, William F. (2021-03-26). "The metabolic network of the last bacterial common ancestor". Communications Biology. 4 (1): 413. doi:10.1038/s42003-021-01918-4. ISSN 2399-3642. PMC 7997952. PMID 33772086.
Other reading
- Wood HG (February 1991). "Life with CO or CO2 and H2 as a source of carbon and energy". FASEB J. 5 (2): 156–63. doi:10.1096/fasebj.5.2.1900793. PMID 1900793. S2CID 45967404.
- Diekert G, Wohlfarth G (1994). "Metabolism of homoacetogens". Antonie van Leeuwenhoek. 66 (1–3): 209–21. doi:10.1007/BF00871640. PMID 7747932. S2CID 7473300.