Resistance-nodulation-cell division superfamily
Resistance-nodulation-division (RND) family transporters are a category of bacterial efflux pumps, especially identified in Gram-negative bacteria and located in the cytoplasmic membrane, that actively transport substrates. The RND superfamily includes seven families: the heavy metal efflux (HME), the hydrophobe/amphiphile efflux-1 (gram-negative bacteria), the nodulation factor exporter family (NFE), the SecDF protein-secretion accessory protein family, the hydrophobe/amphiphile efflux-2 family, the eukaryotic sterol homeostasis family, and the hydrophobe/amphiphile efflux-3 family.[1] These RND systems are involved in maintaining homeostasis of the cell, removal of toxic compounds, and export of virulence determinants.[2] They have a broad substrate spectrum and can lead to the diminished activity of unrelated drug classes if over-expressed. The first reports of drug resistant bacterial infections were reported in the 1940s after the first mass production of antibiotics.[3] Most of the RND superfamily transport systems are made of large polypeptide chains.[4] RND proteins exist primarily in gram-negative bacteria but can also be found in gram-positive bacteria, archaea, and eukaryotes.
| RND permease superfamily | |
|---|---|
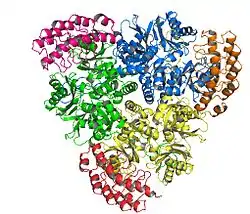 Crystallized AcrB: An HAE-RND subclass protein involved in drug and amphiphilic efflux | |
| Identifiers | |
| Symbol | RND_Permease |
| Pfam clan | CL0322 |
| TCDB | 2.A.6 |
| OPM superfamily | 16 |
| OPM protein | 2gif |
Function
The RND protein dictates the substrate for the completed transport systems including: metal ions, xenobiotics or drugs. Transport of hydrophobic and amphiphilic compounds are carried out by the HAE-RND subfamily. While the efflux of heavy metals are preformed HME-RND.[5]
Mechanism and structure
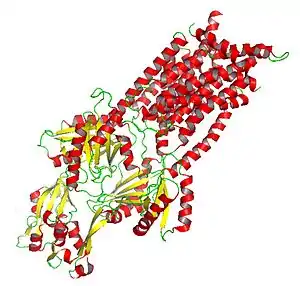
RND proteins are large and can include more than 1000 amino acid residues. They are generally composed of two homologous subunits (suggesting they arose as a result of an intragenic tandem duplication event that occurred in the primordial system prior to divergence of the family members) each containing a periplasmic loop adjacent to 12 transmembrane helices. Of the twelve helices there is a single transmembrane spanner (TMS) at the N-terminus followed by a large extracytoplasmic domain, then six additional TMSs, a second large extracytoplasmic domain, and five final C-terminal TMSs. TM4 governs the specificity for a particular substrate in a given RND protein. Therefore, TM4 can be an indicator for RND specificity without explicit knowledge of the remainder of the protein.[6]
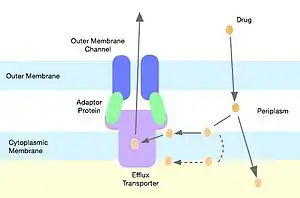
RND pumps are the cytoplasmic residing portion of a complete tripartite complex (Fig. 1) which spreads across the outer-membrane and the inner membrane of gram-negative bacteria, also commonly referred to as the CBA efflux system. The RND protein associates with an outer membrane channel and a periplasmic adaptor protein, and the association of all three proteins allows the system to export substrates into the external medium, providing a huge advantage for the bacteria.[7]
The CusA protein, a HME-RND member transporter, was able to be crystallized providing valuable structural information of HME-RND pumps. CusA exists as a homotrimer with each unit consisting of 12 transmembrane helices (TM1-TM12). The periplasmic domain consists of two helices, TM2 and TM8. In addition, the periplasmic domain is made up of six subdomains, PN1, PN2, PC1, PC2, DN, DC, which form a central pore and a dock domain. The central pore is formed by PN1, PN2, PC1, PC2, and together stabilize the trimeric organization of the homotrimer.[8]
Metal ion efflux (HME-RND)
The HME-RND family functions as the central protein pump in metal ion efflux powered by a proton-substrate antiport. The family includes pumps which export monovalent metals—the Cus system, and pumps which export divalent metals—the Czc system.[5]
Heavy metal resistance by the RND family was first discovered in R. metallidurans through the CzcA and later the CnrA protein. The best characterized RND proteins include CzcCBA (Cd2+, Zn2+, and Co2+), CnrCBA (Ni2+ and Co2+), and NccCBA (Ni2+, Co2+ and Cd2+) in Cupriavidus, Czr (Cd2+ and Zn2+ resistance) in Pseudomonas aeruginosa, and Czn (Cd2+, Zn2+, and Ni2+ resistance) in Helicobacter pylori.[9] It has been proposed that metal-ion efflux occurs from the cytoplasm and periplasm based on the location of multiple substrate binding sites on the RND protein.[6]
CznCBA
The Czn system maintains homeostasis of Cadmium, Zinc, and Nickel resistance; it is involved in Urease modulation, and gastric colonization by H. pylori. The CznC and CznA proteins play the dominating role in nickel homeostasis.[10]
CzcCBA
Czc confers resistance to Cobalt, Zinc, and Cadmium. The CzcCBA operon includes: CzcA (the RND family specific protein), the membrane fusion protein (MFP) CzcB, and the outer membrane factor protein (OMF) CzcC, all of which form the active tripartite complex, and the czcoperon. Expression of the operon is regulated through metal ions.[6]
Drug resistance (HAE-RND)
The RND family plays an important role in producing intrinsic and elevated multi-drug resistance in gram-negative bacteria. The export of amphiphilic and hydrophobic substrates is governed by the HAE-RND family. In E. coli five RND pumps have been specifically identified: AcrAB, AcrAD, AcrEF, MdtEF, and MdtAB. Although it is not clear how the tripartite complex works in bacteria two mechanisms have been proposed: Adaptor Bridging Model and Adaptor Wrapping Model.
HAE-RNDs involvement in the detoxification and exportation of organic substrates allowed for recent characterization of specific pumps due to their increasing medical relevance. Half of the antibiotic resistance demonstrated in in vivo hospital strains of Pseudomonas aeruginosa was attributed to RND efflux proteins. P. aeruginosa contain 13 RND transport systems, including one HME-RND and the remaining HAE-RNDs. Among the best identified are the Mex proteins: MexB, MexD, and MexF, which detoxify organic substances. It is proposed that the MexB systems demonstrates substrate specificity for beta-lactams; while the MexD-system expresses specificity for cepheme compounds.[6]
E. coli – AcrB
In E. coli multi-drug resistance develops from a variety of mechanisms. Particularly concerning is the ability of efflux mechanisms to confer broad-spectrum resistance. RND efflux pumps provide extrusion for a range of compounds. Five protein transporters in E. coli cells that belong to the HAE-RND subfamily have been classified,[11] including the multi-drug efflux protein AcrB, the outer membrane protein TolC and the periplasmic adaptor protein AcrA.[12] The TolC and AcrA proteins are also utilized in the tripartite complex in other identified RND efflux proteins.[11] The AcrAB-TolC efflux system is responsible for the efflux of antimicrobial drugs like penicillin G, cloxacillin, nafcillin, macrolides, novobiocin, linezolid, and fusidic acid antibiotics. Other substrates include dyes, detergents, some organic solvents, and steroid hormones. The ways in which the lipophilic domains of the substrate and the RND pumps is not completely defined.
The crystallized AcrB protein, provides insight into the mechanism of action of HAE-RND proteins, and other RND family proteins.[6]
Multidrug transport (Mdt) efflux
Mdt(A) is an efflux pump that confers resistance to a variety of drugs. It is expressed in L. lactis, E. coli and various other bacteria. Unlike other RND proteins Mdt(A) contains a putative ATP-binding site and two C-motifs conserved in its fifth TMS. Mdt is effective at providing the bacteria with resistance to tetracycline, chloramphenicol, lincosamides and streptomycin. The source of energy for active efflux by Mdt(A) is currently unknown.[13]
References
- Tseng TT, Gratwick KS, Kollman J, Park D, Nies DH, Goffeau A, Saier MH (August 1999). "The RND permease superfamily: an ancient, ubiquitous and diverse family that includes human disease and development proteins". Journal of Molecular Microbiology and Biotechnology. 1 (1): 107–25. PMID 10941792.
- Coyne S, Rosenfeld N, Lambert T, Courvalin P, Périchon B (October 2010). "Overexpression of resistance-nodulation-cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii". Antimicrobial Agents and Chemotherapy. 54 (10): 4389–93. doi:10.1128/AAC.00155-10. PMC 2944555. PMID 20696879.
- Routh, Mathew David, "Structure, function, and regulation of multidrug export proteins among the RND superfamily in Gram-negative bacteria" (2010). Graduate Theses and Dissertations. Paper 11401. http://lib.dr.iastate.edu/etd/11401
- "2.A.6 The Resistance-Nodulation-Cell Division (RND) Superfamily". Transporter Classification Database.
- Moraleda-Muñoz A, Pérez J, Extremera AL, Muñoz-Dorado J (September 2010). "Differential regulation of six heavy metal efflux systems in the response of Myxococcus xanthus to copper". Applied and Environmental Microbiology. 76 (18): 6069–76. Bibcode:2010ApEnM..76.6069M. doi:10.1128/AEM.00753-10. PMC 2937488. PMID 20562277.
- Nies DH (June 2003). "Efflux-mediated heavy metal resistance in prokaryotes". FEMS Microbiology Reviews. 27 (2–3): 313–39. doi:10.1016/s0168-6445(03)00048-2. PMID 12829273.
- Nikaido H (2011). "Structure and mechanism of RND-type multidrug efflux pumps". Advances in Enzymology and Related Areas of Molecular Biology. Advances in Enzymology and Related Areas of Molecular Biology. Vol. 77. pp. 1–60. doi:10.1002/9780470920541.ch1. ISBN 9780470920541. PMC 3122131. PMID 21692366.
- Long F, Su CC, Zimmermann MT, Boyken SE, Rajashankar KR, Jernigan RL, Yu EW (September 2010). "Crystal structures of the CusA efflux pump suggest methionine-mediated metal transport". Nature. 467 (7314): 484–8. Bibcode:2010Natur.467..484L. doi:10.1038/nature09395. PMC 2946090. PMID 20865003.
- Valencia EY, Braz VS, Guzzo C, Marques MV (April 2013). "Two RND proteins involved in heavy metal efflux in Caulobacter crescentus belong to separate clusters within proteobacteria". BMC Microbiology. 13: 79. doi:10.1186/1471-2180-13-79. PMC 3637150. PMID 23578014.
- Stähler FN, Odenbreit S, Haas R, Wilrich J, Van Vliet AH, Kusters JG, Kist M, Bereswill S (July 2006). "The novel Helicobacter pylori CznABC metal efflux pump is required for cadmium, zinc, and nickel resistance, urease modulation, and gastric colonization". Infection and Immunity. 74 (7): 3845–52. doi:10.1128/IAI.02025-05. PMC 1489693. PMID 16790756.
- Anes J, McCusker MP, Fanning S, Martins M (2015). "The ins and outs of RND efflux pumps in Escherichia coli". Frontiers in Microbiology. 6: 587. doi:10.3389/fmicb.2015.00587. PMC 4462101. PMID 26113845.
- Nikaido H, Takatsuka Y (May 2009). "Mechanisms of RND multidrug efflux pumps". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1794 (5): 769–81. doi:10.1016/j.bbapap.2008.10.004. PMC 2696896. PMID 19026770.
- Perreten V, Schwarz FV, Teuber M, Levy SB (April 2001). "Mdt(A), a new efflux protein conferring multiple antibiotic resistance in Lactococcus lactis and Escherichia coli". Antimicrobial Agents and Chemotherapy. 45 (4): 1109–14. doi:10.1128/AAC.45.4.1109-1114.2001. PMC 90432. PMID 11257023.