Retinotopy
Retinotopy (from Greek τόπος, place) is the mapping of visual input from the retina to neurons, particularly those neurons within the visual stream. For clarity, 'retinotopy' can be replaced with 'retinal mapping', and 'retinotopic' with 'retinally mapped'.
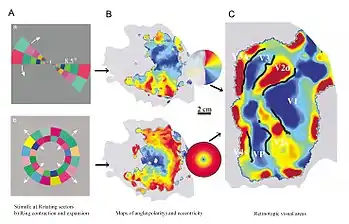
Visual field maps (retinotopic maps) are found in many amphibian and mammalian species, though the specific size, number, and spatial arrangement of these maps can differ considerably. Sensory topographies can be found throughout the brain and are critical to the understanding of one's external environment. Moreover, the study of sensory topographies and retinotopy in particular has furthered our understanding of how neurons encode and organize sensory signals.
Retinal mapping of the visual field is maintained through various points of the visual pathway including but not limited to the retina, the dorsal lateral geniculate nucleus, the optic tectum, the primary visual cortex (V1), and higher visual areas (V2-V4). Retinotopic maps in cortical areas other than V1 are typically more complex, in the sense that adjacent points of the visual field are not always represented in adjacent regions of the same area. For example, in the second visual area (V2), the map is divided along an imaginary horizontal line across the visual field, in such a way that the parts of the retina that respond to the upper half of the visual field are represented in cortical tissue that is separated from those parts that respond the lower half of the visual field. Even more complex maps exist in the third and fourth visual areas V3 and V4, and in the dorsomedial area (V6). In general, these complex maps are referred to as second-order representations of the visual field, as opposed to first-order (continuous) representations such as V1.[1] Additional retinotopic regions include ventral occipital (VO-1, VO-2),[2] lateral occipital (LO-1, LO-2),[3] dorsal occipital (V3A, V3B),[4] and posterior parietal cortex (IPS0, IPS1, IPS2, IPS3, IPS4).[5]
History
The discovery of visual field maps in humans can be traced to neurological cases, arising from war injuries, described and analyzed independently by Tatsuji Inouye (a Japanese ophthalmologist) and Gordon Holmes (a British neurologist). They observed correlations between the position of the entry wound and visual field loss (see Fishman, 1997[6] for an historical review).
Development
Molecular Cues
The "Chemoaffinity Hypothesis" was established by Sperry et al in 1963 in which it is thought that molecular gradients in both presynaptic and postsynaptic partners within the optic tectum organize developing axons into a coarse retinotopic map.[7] This was established after a series of seminal experiments in fish and amphibians showed that retinal ganglion axons were already retinotopically organized within the optic tract and if severed, would regenerate and project back to retinotopically appropriate locations. Later, it was identified that receptor tyrosine kinases family EphA and a related EphA binding molecule referred to as ephrin-A family are expressed in complementary gradients in both the retina and the tectum.[8][9][10] More specifically in the mouse, Ephrin A5 is expressed along the rostral-caudal axis of the optic tectum[11] whereas the EphB family is expressed along the medio-lateral axis.[12] This bimodal expression suggests a mechanism for the graded mapping of the temporal-nasal axis and the dorsoventral axis of the retina.
Target Space
While molecular cues are thought to guide axons into a coarse retinotopic map, the resolution of this map is thought to be influenced by available target space on postsynaptic partners. In wild type mice, it is thought that competition of target space is important for ensuring continuous retinal mapping, and that if perturbed, this competition may lead to the expansion or compression of the map depending on the available space. If the available space is altered, such as lesioning or ablating half of the retina, the healthy axons will expand their arbors in the tectum to fill the space.[13] Similarly, if part of the tectum is ablated, the retinal axons will compress the topography to fit within the available tectal space.[14]
Neural Activity
While neural activity in the retina is not necessary for the development of retinotopy, it seems to be a critical component for the refinement and stabilization of connectivity. Dark reared animals (no external visual cues) develop a normal retinal map in the tectum with no marked changes in receptive field size or laminar organization.[15][16] While these animals may not have received external visual cues during development, these experiments suggest that spontaneous activity in the retina may be sufficient for retinotopic organization. In the goldfish, no neural activity (no external visual cues, and no spontaneous activity) did not prevent the formation of the retinal map but the final organization showed signs of lower resolution refinement and more dynamic growth (less stable).[17] Based on Hebbian mechanisms, the thought is that if neurons are sensitive to similar stimuli (similar area of the visual field, similar orientation or direction selectivity) they will likely fire together. This patterned firing will result in stronger connectivity within the retinotopic organization through NMDAR synapse stabilization mechanisms in the post synaptic cells.[18][19]
Dynamic Growth
Another important factor in the development of retinotopy is the potential for structural plasticity even after neurons are morphologically mature. One interesting hypothesis is that axons and dendrites are continuously extending and retracting their axons and dendrites. Several factors alter this dynamic growth including the Chemoaffinity Hypothesis, the presence of developed synapses, and neural activity. As the nervous system develops and more cells are added, this structural plasticity allows for axons to gradually refine their place within the retinotopy.[20] This plasticity is not specific to retinal ganglion axons, rather it's been shown that dendritic arbors of tectal neurons and filopodial processes of radial glial cells are also highly dynamic.
Description
In many locations within the brain, adjacent neurons have receptive fields that include slightly different, but overlapping portions of the visual field. The position of the center of these receptive fields forms an orderly sampling mosaic that covers a portion of the visual field. Because of this orderly arrangement, which emerges from the spatial specificity of connections between neurons in different parts of the visual system, cells in each structure can be seen as contributing to a map of the visual field (also called a retinotopic map, or a visuotopic map). Retinotopic maps are a particular case of topographic organization. Many brain structures that are responsive to visual input, including much of the visual cortex and visual nuclei of the brain stem (such as the superior colliculus) and thalamus (such as the lateral geniculate nucleus and the pulvinar), are organized into retinotopic maps, also called visual field maps.
Areas of the visual cortex are sometimes defined by their retinotopic boundaries, using a criterion that states that each area should contain a complete map of the visual field. However, in practice the application of this criterion is in many cases difficult.[1] Those visual areas of the brainstem and cortex that perform the first steps of processing the retinal image tend to be organized according to very precise retinotopic maps. The role of retinotopy in other areas, where neurons have large receptive fields, is still being investigated.[21]
.jpg.webp)
Retinotopy mapping shapes the folding of the cerebral cortex. In both the V1 and V2 areas of macaques and humans the vertical meridian of their visual field tends to be represented on the cerebral cortex's convex gyri folds whereas the horizontal meridian tends to be represented in their concave sulci folds.[22]
Methods
Retinotopy mapping in humans is done with functional Magnetic Resonance Imaging (fMRI). The subject inside the fMRI machine focuses on a point. Then the retina is stimulated with a circular image or angled lines about focus point.[23][24][25] The radial map displays the distance from the center of vision. The angular map shows angular location using rays angled about the center of vision. Combining the radial and angular maps, you can see the separate regions of the visual cortex and the smaller maps in each region.
See also
References
- Rosa MG (December 2002). "Visual maps in the adult primate cerebral cortex: some implications for brain development and evolution". Brazilian Journal of Medical and Biological Research. 35 (12): 1485–98. doi:10.1590/s0100-879x2002001200008. PMID 12436190.
- Brewer AA, Liu J, Wade AR, Wandell BA (August 2005). "Visual field maps and stimulus selectivity in human ventral occipital cortex". Nature Neuroscience. 8 (8): 1102–9. doi:10.1038/nn1507. PMID 16025108. S2CID 8413534.
- Larsson J, Heeger DJ (December 2006). "Two retinotopic visual areas in human lateral occipital cortex". The Journal of Neuroscience. 26 (51): 13128–42. doi:10.1523/jneurosci.1657-06.2006. PMC 1904390. PMID 17182764.
- Tootell RB, Mendola JD, Hadjikhani NK, Ledden PJ, Liu AK, Reppas JB, Sereno MI, Dale AM (September 1997). "Functional analysis of V3A and related areas in human visual cortex". The Journal of Neuroscience. 17 (18): 7060–78. doi:10.1523/JNEUROSCI.17-18-07060.1997. PMC 6573277. PMID 9278542.
- Silver MA, Ress D, Heeger DJ (August 2005). "Topographic maps of visual spatial attention in human parietal cortex". Journal of Neurophysiology. 94 (2): 1358–71. doi:10.1152/jn.01316.2004. PMC 2367310. PMID 15817643.
- Ronald S. Fishman (1997). Gordon Holmes, the cortical retina, and the wounds of war. The seventh Charles B. Snyder Lecture Documenta Ophthalmologica 93: 9-28, 1997.
- Sperry, R. W. (October 1963). "Chemoaffinity in the Orderly Growth of Nerve Fiber Patterns and Connections". Proceedings of the National Academy of Sciences of the United States of America. 50 (4): 703–710. Bibcode:1963PNAS...50..703S. doi:10.1073/pnas.50.4.703. ISSN 0027-8424. PMC 221249. PMID 14077501.
- Drescher, Uwe; Kremoser, Claus; Handwerker, Claudia; Löschinger, Jürgen; Noda, Masaharu; Bonhoeffer, Friedrich (1995-08-11). "In vitro guidance of retinal ganglion cell axons by RAGS, a 25 kDa tectal protein related to ligands for Eph receptor tyrosine kinases". Cell. 82 (3): 359–370. doi:10.1016/0092-8674(95)90425-5. ISSN 0092-8674. PMID 7634326. S2CID 2537692.
- Feldheim, David A.; O'Leary, Dennis D. M. (November 2010). "Visual map development: bidirectional signaling, bifunctional guidance molecules, and competition". Cold Spring Harbor Perspectives in Biology. 2 (11): a001768. doi:10.1101/cshperspect.a001768. ISSN 1943-0264. PMC 2964178. PMID 20880989.
- Brennan, C.; Monschau, B.; Lindberg, R.; Guthrie, B.; Drescher, U.; Bonhoeffer, F.; Holder, N. (February 1997). "Two Eph receptor tyrosine kinase ligands control axon growth and may be involved in the creation of the retinotectal map in the zebrafish". Development. 124 (3): 655–664. doi:10.1242/dev.124.3.655. ISSN 0950-1991. PMID 9043080.
- Suetterlin, Philipp; Drescher, Uwe (2014-11-19). "Target-independent ephrina/EphA-mediated axon-axon repulsion as a novel element in retinocollicular mapping". Neuron. 84 (4): 740–752. doi:10.1016/j.neuron.2014.09.023. ISSN 1097-4199. PMC 4250266. PMID 25451192.
- Hindges, Robert; McLaughlin, Todd; Genoud, Nicolas; Henkemeyer, Mark; O'Leary, Dennis D. M. (2002-08-01). "EphB forward signaling controls directional branch extension and arborization required for dorsal-ventral retinotopic mapping". Neuron. 35 (3): 475–487. doi:10.1016/s0896-6273(02)00799-7. ISSN 0896-6273. PMID 12165470. S2CID 18724075.
- Schmidt, J. T.; Easter, S. S. (1978-02-15). "Independent biaxial reorganization of the retinotectal projection: a reassessment". Experimental Brain Research. 31 (2): 155–162. doi:10.1007/BF00237596. hdl:2027.42/46548. ISSN 0014-4819. PMID 631237. S2CID 8865051.
- Yoon, M. G. (June 1976). "Progress of topographic regulation of the visual projection in the halved optic tectum of adult goldfish". The Journal of Physiology. 257 (3): 621–643. doi:10.1113/jphysiol.1976.sp011388. ISSN 0022-3751. PMC 1309382. PMID 950607.
- Keating, M. J.; Grant, S.; Dawes, E. A.; Nanchahal, K. (February 1986). "Visual deprivation and the maturation of the retinotectal projection in Xenopus laevis". Journal of Embryology and Experimental Morphology. 91: 101–115. ISSN 0022-0752. PMID 3711779.
- Nevin, Linda M; Taylor, Michael R; Baier, Herwig (2008-12-16). "Hardwiring of fine synaptic layers in the zebrafish visual pathway". Neural Development. 3: 36. doi:10.1186/1749-8104-3-36. ISSN 1749-8104. PMC 2647910. PMID 19087349.
- Meyer, R. L. (February 1983). "Tetrodotoxin inhibits the formation of refined retinotopography in goldfish". Brain Research. 282 (3): 293–298. doi:10.1016/0165-3806(83)90068-8. ISSN 0006-8993. PMID 6831250.
- Rajan, I.; Witte, S.; Cline, H. T. (1999-02-15). "NMDA receptor activity stabilizes presynaptic retinotectal axons and postsynaptic optic tectal cell dendrites in vivo". Journal of Neurobiology. 38 (3): 357–68. doi:10.1002/(SICI)1097-4695(19990215)38:3<357::AID-NEU5>3.0.CO;2-#. ISSN 0022-3034. PMID 10022578.
- Schmidt, J. T.; Buzzard, M.; Borress, R.; Dhillon, S. (2000-02-15). "MK801 increases retinotectal arbor size in developing zebrafish without affecting kinetics of branch elimination and addition". Journal of Neurobiology. 42 (3): 303–314. doi:10.1002/(SICI)1097-4695(20000215)42:3<303::AID-NEU2>3.0.CO;2-A. ISSN 0022-3034. PMID 10645970.
- Sakaguchi, D. S.; Murphey, R. K. (December 1985). "Map formation in the developing Xenopus retinotectal system: an examination of ganglion cell terminal arborizations". The Journal of Neuroscience. 5 (12): 3228–3245. doi:10.1523/JNEUROSCI.05-12-03228.1985. ISSN 0270-6474. PMC 6565231. PMID 3001241.
- Wandell BA, Brewer AA, Dougherty RF (April 2005). "Visual field map clusters in human cortex". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 360 (1456): 693–707. doi:10.1098/rstb.2005.1628. PMC 1569486. PMID 15937008.
- Rajimehr R, Tootell RB (September 2009). "Does retinotopy influence cortical folding in primate visual cortex?". The Journal of Neuroscience. 29 (36): 11149–52. doi:10.1523/JNEUROSCI.1835-09.2009. PMC 2785715. PMID 19741121.
- Bridge H (March 2011). "Mapping the visual brain: how and why". Eye. 25 (3): 291–6. doi:10.1038/eye.2010.166. PMC 3178304. PMID 21102491.>
- DeYoe EA, Carman GJ, Bandettini P, Glickman S, Wieser J, Cox R, Miller D, Neitz J (March 1996). "Mapping striate and extrastriate visual areas in human cerebral cortex". Proceedings of the National Academy of Sciences of the United States of America. 93 (6): 2382–6. Bibcode:1996PNAS...93.2382D. doi:10.1073/pnas.93.6.2382. PMC 39805. PMID 8637882.
- Engel SA, Glover GH, Wandell BA (March 1997). "Retinotopic organization in human visual cortex and the spatial precision of functional MRI". Cerebral Cortex. 7 (2): 181–92. doi:10.1093/cercor/7.2.181. PMID 9087826.