SKIV2L
Helicase SKI2W is an enzyme that in humans is encoded by the SKIV2L gene.[5][6][7] This enzyme is a human homologue of yeast SKI2, which may be involved in antiviral activity by blocking translation of poly(A) deficient mRNAs. The SKIV2L gene is located in the class III region of the major histocompatibility complex.[6]
| SKIV2L | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | SKIV2L, 170A, DDX13, HLP, SKI2, SKI2W, SKIV2, THES2, SKIV2L1, Ski2 like RNA helicase | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 600478 MGI: 1099835 HomoloGene: 123971 GeneCards: SKIV2L | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


DEAD box proteins, characterized by the conserved motif Asp-Glu-Ala-Asp (DEAD), are putative RNA helicases. They are implicated in a number of cellular processes involving alteration of RNA secondary structure such as translation initiation, nuclear and mitochondrial splicing, and ribosome and spliceosome assembly. Based on their distribution patterns, some members of this family are believed to be involved in embryogenesis, spermatogenesis, and cellular growth and division.
Identification of gene
A novel human cDNA, homologous to the yeast gene SKI2, was identified in 1995. Researchers localised the corresponding gene to chromosome 6p.21.[8]
The SKIV2L gene, also known as SKI2W or Ski2-like RNA Helicase, spans 11Kb and contains 28 exons.[7] Located between RD and RP1 gene in MHC III on the short arm of chromosome 6, it has 16 transcripts on Ensembl, 3 of which are protein coding. One of these proteins, SKI2W, has 1246 amino acids and a helicase binding domain between amino acids 319-475, and is thought to be involved in exosome RNA mediated RNA decay.[9]
The presence of the DEVH-box at position 423-426 within SKI2W infers that it is a member of the SF-II helicase family.
The human protein was named SKI2W because of its similarity to yeast protein Ski2, which has highly homologous (nearly identical) regions at the helicase domain and the C terminal.[10]
SKIV2L transcripts are expressed in most, if not all, human tissue tested including spleen, thymus, small intestine, colon, heart, brain and liver.[11]
| Species | Gene | Locus | CCDS code | UniProt | Exons | Amino acids |
|---|---|---|---|---|---|---|
| Human | SKIV2L | 6p21.33 | 4731.1 | Q15477 | 28 | 1246 |
| Mouse | Skiv2l | chr 17 | 28661.1 | n/a | 28 | 1244 |
Function
The majority of the eukaryotic genome is transcribed into RNA molecules, which generates pools of RNA that require processing and surveillance in order to control abundant and damaged material. The RNA exosome multiprotein complex performs this function and is dependent on cofactors. The exosome was initially discovered in yeast but is also present in higher eukaryotes. It has activity in both the nucleus and cytoplasm for normal mRNA decay and for RNA surveillance and quality control through nonsense mediated; non-stop and no-go decay.[12]
.png.webp)
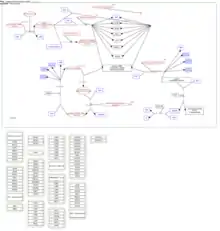
SKI2W is part of the tetraprotein ski complex which is an obligatory cytoplasmic cofactor of the RNA exosome and consists of SKI2W, TTC37 and 2 subunits of WD40 (encoded by WDR61), as pictured in Figure 1.[13] Much of the information on SKI2W function is from yeast studies, where the homologue for SKI2W is ski2. In yeast, ski2 forms a ski complex with ski3 and 2 subunits of ski8. The ski2 (the SKI2W homolog in yeast) contains DEVH-box proteins which suggests it is the only protein in the ski complex to have an enzymatic helicase function.[14] The exact interactions are not well described, however DEVH-box helicases are shown to separate nucleic strands in an energy dependent manner. The yeast ski complex has been more extensively studied than the human homologue, and a crystal structure of the RNA exosome and its interactions has been created that supports a role in non-stop decay, to thereby protect the cell from aberrant proteins.[14]
Ski2 protein in yeast is also thought to have a role in antiviral defence, probably via its role in RNA turnover or through control of RNA degradation.[16]
SKIV2L has been shown to be a negative regulator of the Rig-I like receptors (RLRs) that detect RNA. The authors found that the cytosolic RNA exosome, defined by the SKIV2L RNA helicase, is important for limiting the activation of RLRs and the antiviral response. If the endogenous RNAs fail to be processed, the cell undergoes an unfolded protein response which triggers an antiviral interferon (IFN) response. Human cells with SKIV2L deficiency are shown to have a strong IFN signature suggestive of a chronic antiviral response. The authors suggest that patients could subsequently be more prone to autoimmune disorders, although this has not been shown as yet, possibly due to the high mortality of patients. However this finding does suggest a link between SKIV2L and immune response.[17]
Clinical significance
Pathogenic variants in SKIV2L has been linked to Tricho-hepato-enteric syndrome (THES), also known as syndromic diarrhoea (SD) or phenotypic diarrhoea (PD). First described by Stankler et al (1982) as Stankler syndrome, this condition was renamed THES in 1994.[18] THES is rare, with an estimated prevalence of 1:1,000,000.[19] It is characterised by intractable diarrhoea, beginning in first few weeks of life; characteristic hair abnormalities, ”woolly" and brittle hair, intrauterine growth restriction and characteristic facial dysmorphisms. Other associations are hepatic dysfunction, skin abnormalities, intellectual disability and immunodeficiency. Less common findings include platelet abnormalities and congenital heart defects.[20]
There are two causative genes, SKIV2L (in 1/3 of patients) and TTC37 (2/3 of patients), both encode for proteins in the Ski complex and clinically they are indistinguishable from each other.[12]
Inherited in an autosomal recessive manner with complete penetrance, approximately 2/3rd of patients are homozygotes and 1/3rd are compound heterozygous. Mutations are spread throughout the gene with no identifiable hotspot and generally consist of frameshift, missense and nonsense mutations; a smaller number are splice site mutations.[13] There is no clear genotype/phenotype correlation with overall disease severity, even siblings with the same homozygous mutation display variable phenotypes. Patients have been described globally in Europe, Saudi Arabia, Malaysia, China and Japan.[21][22][10][23]
Intractable watery diarrhoea is a near consistent feature in almost all described cases, almost always starting soon after birth and commonly requiring parenteral nutrition. In cases which do not require parenteral nutrition an elemental diet and supplemental feeding is required.[13]
The majority of SKIV2L pathogenic variant harbouring patients are small at birth (<10th centile) and remain growth restricted despite increased nutrition. Hair abnormalities are seen in >90% of patients and are described as woolly, brittle hair that is easily removed.[13]
Facial dysmorphisms are found in most patients and become more apparent with age. These include a large forehead, broad base of the nose and hypertelorism. Overall the facial features are described as “coarse”.
Liver disease is reported frequently in SKIV2L patients (>80%), ranging from fibrosis, cirrhosis, hepatomegaly and raised liver enzymes. Histopathology when performed shows iron overload and can be consistent with haemochromatosis.[19]
Skin abnormalities are frequently reported and are variable including café au lait lesions, haemangiomas and xerosis. A report from Saudi Arabia suggested the skin changes were more frequent in the lower limb and pelvic region of their regional cohort.[22]
Immunodeficiency is reported in some patients. It is poorly delineated and mainly consists of low immunoglobulins and inadequate vaccine responses, however hyper IgA has also been reported. Immunoglobulin therapy has been shown to lower rates of infection.[24]
Less commonly congenital cardiac defects have been reported, mostly ventricular septal defects (VSD), atrial septal defects (ASD), and rarely Tetralogy of Fallot and peripheral pulmonary stenosis.
Mortality rate
Initially reported as high as 62.5% with most deaths in the first year, more recent reports have estimated mortality of around 30%, which is similar to other disorders which are dependent on parenteral nutrition.
Recommended treatment and surveillance
No specific treatments are available for THES. The goal is to maximise weight gain and reduce infection rates.
Most children require parenteral nutrition (PN) which can be combined with oral feeding, most commonly a semi-elemental diet which allows patients to become independent of PN over time. Nutrition and growth should be closely monitored. If PN is not required, reports have described the use of an amino acid based formula, although it is unclear if weight gain was adequate.
Immunoglobulin levels and vaccine responses should be tested. If any abnormalities are found then a paediatric immunologist should be consulted and intravenous immunoglobulin (IVIG) can be considered to reduce chance of systemic infections.[20][25] Infection was reported as a cause of death in 20% of a large cohort of French patients.[13]
A recent study looked at immunodeficiency in 9 THES patients, of which 3 had SKIV2L pathogenic variants. The authors reported that the degranulation and number of IFN-γ producing NK cells were reduced in most patients (although it is unclear if this included the SKIV2L patients) and proposed that this could lead to susceptibility to RNA viruses, with 4/9 patients harbouring a chronic EBV infection and one patient dying of measles.[24]
Regular liver assessment should include ultrasound and hepatic enzymes and developmental assessment should be performed.[26]
Steroids, immunosuppressants and haematopoietic stem cell transplant has been THES with no success and therefore it is not recommended.
Genetic counselling should be offered as a sibling recurrence is 25% with each conception.
Age related macular degeneration (ARMD)
An intronic single nucleotide polymorphism (SNP) in SKIV2L on genome wide association studies has been shown to be protective for age related macular degeneration. 3’UTR variant in SKIV2L has recently been reported to exert a protective effect in polypoidal choroidal vasculopathy, a haemorrhagic macular disease that shares some features with neovascular ARMD. As the variants would not affect structure of the protein, it was proposed that it affects regulation of oxidative stress pathways.[27]
Conversely another study showed a genetic variant rs429608 to be strongly associated with the development of ARMD in the Han Chinese population,[28] however further studies are needed to investigate the biological role of SKIV2L and pathogenesis.
Notes
References
- ENSG00000204351, ENSG00000228896, ENSG00000206353, ENSG00000223493, ENSG00000225737 GRCh38: Ensembl release 89: ENSG00000232616, ENSG00000204351, ENSG00000228896, ENSG00000206353, ENSG00000223493, ENSG00000225737 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000040356 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Lee SG, Lee I, Park SH, Kang C, Song K (February 1995). "Identification and characterization of a human cDNA homologous to yeast SKI2". Genomics. 25 (3): 660–6. doi:10.1016/0888-7543(95)80008-A. PMID 7759100.
- "SKIV2L superkiller viralicidic activity 2-like (S. cerevisiae)". Entrez Gene. National Center for Biotechnology Information, U.S. National Library of Medicine.
- Yang Z, Shen L, Dangel AW, Wu LC, Yu CY (November 1998). "Four ubiquitously expressed genes, RD (D6S45)-SKI2W (SKIV2L)-DOM3Z-RP1 (D6S60E), are present between complement component genes factor B and C4 in the class III region of the HLA". Genomics. 53 (3): 338–47. doi:10.1006/geno.1998.5499. PMID 9799600.
- Lee SG, Lee I, Park SH, Kang C, Song K (February 1995). "Identification and characterization of a human cDNA homologous to yeast SKI2". Genomics. 25 (3): 660–6. doi:10.1016/0888-7543(95)80008-a. PMID 7759100.
- "UniProt: the universal protein knowledgebase". Nucleic Acids Research. 45 (D1): D158–D169. January 2017. doi:10.1093/nar/gkw1099. PMC 5210571. PMID 27899622.
- Lee WS, Teo KM, Ng RT, Chong SY, Kee BP, Chua KH (July 2016). "Novel mutations in SKIV2L and TTC37 genes in Malaysian children with trichohepatoenteric syndrome". Gene. 586 (1): 1–6. doi:10.1016/j.gene.2016.03.049. PMID 27050310.
- Qu X, Yang Z, Zhang S, Shen L, Dangel AW, Hughes JH, et al. (September 1998). "The human DEVH-box protein Ski2w from the HLA is localized in nucleoli and ribosomes". Nucleic Acids Research. 26 (17): 4068–77. doi:10.1093/nar/26.17.4068. PMC 147813. PMID 9705521.
- Fabre A, Charroux B, Martinez-Vinson C, Roquelaure B, Odul E, Sayar E, et al. (April 2012). "SKIV2L mutations cause syndromic diarrhea, or trichohepatoenteric syndrome". American Journal of Human Genetics. 90 (4): 689–92. doi:10.1016/j.ajhg.2012.02.009. PMC 3322239. PMID 22444670.
- Bourgeois P, Esteve C, Chaix C, Béroud C, Lévy N, Fabre A, Badens C (June 2018). "Tricho-Hepato-Enteric Syndrome mutation update: Mutations spectrum of TTC37 and SKIV2L, clinical analysis and future prospects". Human Mutation. 39 (6): 774–789. doi:10.1002/humu.23418. PMID 29527791. S2CID 4331400.
- Halbach F, Reichelt P, Rode M, Conti E (August 2013). "The yeast ski complex: crystal structure and RNA channeling to the exosome complex". Cell. 154 (4): 814–26. doi:10.1016/j.cell.2013.07.017. PMID 23953113.
- "Deadenylation-dependent mRNA decay (Homo sapiens) - WikiPathways". www.wikipathways.org. Retrieved 2018-12-18.
- Schmidt C, Kowalinski E, Shanmuganathan V, Defenouillère Q, Braunger K, Heuer A, et al. (December 2016). "The cryo-EM structure of a ribosome-Ski2-Ski3-Ski8 helicase complex". Science. 354 (6318): 1431–1433. Bibcode:2016Sci...354.1431S. doi:10.1126/science.aaf7520. PMID 27980209. S2CID 30262907.
- Eckard SC, Rice GI, Fabre A, Badens C, Gray EE, Hartley JL, et al. (September 2014). "The SKIV2L RNA exosome limits activation of the RIG-I-like receptors". Nature Immunology. 15 (9): 839–45. doi:10.1038/ni.2948. PMC 4139417. PMID 25064072.
- Girault D, Goulet O, Le Deist F, Brousse N, Colomb V, Césarini JP, et al. (July 1994). "Intractable infant diarrhea associated with phenotypic abnormalities and immunodeficiency". The Journal of Pediatrics. 125 (1): 36–42. doi:10.1016/s0022-3476(94)70118-0. PMID 8021782.
- Fabre A, Breton A, Coste ME, Colomb V, Dubern B, Lachaux A, et al. (January 2014). "Syndromic (phenotypic) diarrhoea of infancy/tricho-hepato-enteric syndrome". Archives of Disease in Childhood. 99 (1): 35–8. doi:10.1136/archdischild-2013-304016. PMID 24108068. S2CID 206855850.
- Fabre A, Martinez-Vinson C, Goulet O, Badens C (January 2013). "Syndromic diarrhea/Tricho-hepato-enteric syndrome". Orphanet Journal of Rare Diseases. 8 (1): 5. doi:10.1186/1750-1172-8-5. PMC 3560276. PMID 23302111.
- Hiejima E, Yasumi T, Nakase H, Matsuura M, Honzawa Y, Higuchi H, et al. (November 2017). "Tricho-hepato-enteric syndrome with novel SKIV2L gene mutations: A case report". Medicine. 96 (46): e8601. doi:10.1097/MD.0000000000008601. PMC 5704822. PMID 29145277.
- Monies DM, Rahbeeni Z, Abouelhoda M, Naim EA, Al-Younes B, Meyer BF, Al-Mehaidib A (March 2015). "Expanding phenotypic and allelic heterogeneity of tricho-hepato-enteric syndrome". Journal of Pediatric Gastroenterology and Nutrition. 60 (3): 352–6. doi:10.1097/MPG.0000000000000627. PMID 25714577. S2CID 31110518.
- Zheng B, Pan J, Jin Y, Wang C, Liu Z (September 2016). "Targeted next-generation sequencing identification of a novel missense mutation of the SKIV2L gene in a patient with trichohepatoenteric syndrome". Molecular Medicine Reports. 14 (3): 2107–10. doi:10.3892/mmr.2016.5503. PMID 27431780.
- Vély F, Barlogis V, Marinier E, Coste ME, Dubern B, Dugelay E, et al. (2018-05-11). "Combined Immunodeficiency in Patients With Trichohepatoenteric Syndrome". Frontiers in Immunology. 9: 1036. doi:10.3389/fimmu.2018.01036. PMC 5958188. PMID 29868001.
- Rider NL, Boisson B, Jyonouchi S, Hanson EP, Rosenzweig SD, Casanova JL, Orange JS (2015). "Corrigendum: Novel TTC37 Mutations in a Patient with Immunodeficiency without Diarrhea: Extending the Phenotype of Trichohepatoenteric Syndrome". Frontiers in Pediatrics. 3: 28. doi:10.3389/fped.2015.00028. PMC 4398912. PMID 25932458.
- Fabre A, Bourgeois P, Chaix C, Bertaux K, Goulet O, Badens C (January 2018). "Trichohepatoenteric Syndrome". In Adam MP, Ardinger HH, Pagon RA, Wallace SE, Bean LJ, Stephens K, Amemiya A (eds.). GeneReviews. Seattle: University of Washington. PMID 29334452.
- Kopplin LJ, Igo RP, Wang Y, Sivakumaran TA, Hagstrom SA, Peachey NS, et al. (December 2010). "Genome-wide association identifies SKIV2L and MYRIP as protective factors for age-related macular degeneration". Genes and Immunity. 11 (8): 609–21. doi:10.1038/gene.2010.39. PMC 3375062. PMID 20861866.
- Lu F, Shi Y, Qu C, Zhao P, Liu X, Gong B, et al. (April 2013). "A genetic variant in the SKIV2L gene is significantly associated with age-related macular degeneration in a Han Chinese population". Investigative Ophthalmology & Visual Science. 54 (4): 2911–7. doi:10.1167/iovs.12-11381. PMID 23557739.
Further reading
- Yang Z, Qu X, Yu CY (August 2001). "Features of the two gene pairs RD-SKI2W and DOM3Z-RP1 located between complement component genes factor B and C4 at the MHC class III region". Frontiers in Bioscience. 6: D927-35. doi:10.2741/Yang. PMID 11487501.
- Dangel AW, Shen L, Mendoza AR, Wu LC, Yu CY (June 1995). "Human helicase gene SKI2W in the HLA class III region exhibits striking structural similarities to the yeast antiviral gene SKI2 and to the human gene KIAA0052: emergence of a new gene family". Nucleic Acids Research. 23 (12): 2120–6. doi:10.1093/nar/23.12.2120. PMC 306998. PMID 7610041.
- Shen L, Wu LC, Sanlioglu S, Chen R, Mendoza AR, Dangel AW, Carroll MC, Zipf WB, Yu CY (March 1994). "Structure and genetics of the partially duplicated gene RP located immediately upstream of the complement C4A and the C4B genes in the HLA class III region. Molecular cloning, exon-intron structure, composite retroposon, and breakpoint of gene duplication". The Journal of Biological Chemistry. 269 (11): 8466–76. doi:10.1016/S0021-9258(17)37217-4. PMID 8132574.
- Albertella MR, Jones H, Thomson W, Olavesen MG, Campbell RD (September 1996). "Localization of eight additional genes in the human major histocompatibility complex, including the gene encoding the casein kinase II beta subunit (CSNK2B)". Genomics. 36 (2): 240–51. doi:10.1006/geno.1996.0459. PMID 8812450.
- Lee SG, Song K (June 1997). "Genomic organization of the human DDX13 gene located between RD and RP1 in the class III MHC complex". Molecules and Cells. 7 (3): 414–8. PMID 9264031.
- Qu X, Yang Z, Zhang S, Shen L, Dangel AW, Hughes JH, Redman KL, Wu LC, Yu CY (September 1998). "The human DEVH-box protein Ski2w from the HLA is localized in nucleoli and ribosomes". Nucleic Acids Research. 26 (17): 4068–77. doi:10.1093/nar/26.17.4068. PMC 147813. PMID 9705521.
- Lee SG, Song K (February 2000). "Identification and characterization of a bidirectional promoter from the intergenic region between the human DDX13 and RD genes". Molecules and Cells. 10 (1): 47–53. doi:10.1007/s10059-000-0047-z. PMID 10774746. S2CID 31586458.
- Chen CY, Gherzi R, Ong SE, Chan EL, Raijmakers R, Pruijn GJ, Stoecklin G, Moroni C, Mann M, Karin M (November 2001). "AU binding proteins recruit the exosome to degrade ARE-containing mRNAs". Cell. 107 (4): 451–64. doi:10.1016/S0092-8674(01)00578-5. PMID 11719186. S2CID 14817671.
- Lehner B, Sanderson CM (July 2004). "A protein interaction framework for human mRNA degradation". Genome Research. 14 (7): 1315–23. doi:10.1101/gr.2122004. PMC 442147. PMID 15231747.
- Beausoleil SA, Jedrychowski M, Schwartz D, Elias JE, Villén J, Li J, Cohn MA, Cantley LC, Gygi SP (August 2004). "Large-scale characterization of HeLa cell nuclear phosphoproteins". Proceedings of the National Academy of Sciences of the United States of America. 101 (33): 12130–5. Bibcode:2004PNAS..10112130B. doi:10.1073/pnas.0404720101. PMC 514446. PMID 15302935.
- Beausoleil SA, Villén J, Gerber SA, Rush J, Gygi SP (October 2006). "A probability-based approach for high-throughput protein phosphorylation analysis and site localization". Nature Biotechnology. 24 (10): 1285–92. doi:10.1038/nbt1240. PMID 16964243. S2CID 14294292.
- Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (November 2006). "Global, in vivo, and site-specific phosphorylation dynamics in signaling networks". Cell. 127 (3): 635–48. doi:10.1016/j.cell.2006.09.026. PMID 17081983. S2CID 7827573.
- Yang CT, Hindes AE, Hultman KA, Johnson SL (June 2007). "Mutations in gfpt1 and skiv2l2 cause distinct stage-specific defects in larval melanocyte regeneration in zebrafish". PLOS Genetics. 3 (6): e88. doi:10.1371/journal.pgen.0030088. PMC 1885281. PMID 17542649.