Sarcoscypha occidentalis
Sarcoscypha occidentalis, commonly known as the stalked scarlet cup or the western scarlet cup, is a species of fungus in the family Sarcoscyphaceae of the Pezizales order. Fruit bodies have small, bright red cups up to 2 cm (0.8 in) wide atop a slender whitish stem that is between 1 and 3 cm (0.4 and 1.2 in) long. A saprobic species, it is found growing on hardwood twigs, particularly those that are partially buried in moist and shaded humus-rich soil. The fungus is distributed in the continental United States east of the Rocky Mountains, Central America, the Caribbean, and Asia. It is distinguished from the related species S. coccinea and S. austriaca by differences in geographical distribution, fruiting season, and fruit body structure. Phylogenetic analysis has shown that it is most closely related to other Sarcoscypha species that contain large oil droplets in their spores. The species Molliardiomyces occidentalis is an imperfect form of the fungus that lacks a sexually reproductive stage in its life cycle.
| Sarcoscypha occidentalis | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Fungi |
| Division: | Ascomycota |
| Class: | Pezizomycetes |
| Order: | Pezizales |
| Family: | Sarcoscyphaceae |
| Genus: | Sarcoscypha |
| Species: | S. occidentalis |
| Binomial name | |
| Sarcoscypha occidentalis | |
| Synonyms[1] | |
| Sarcoscypha occidentalis | |
|---|---|
| Smooth hymenium | |
| No distinct cap | |
| Hymenium attachment is irregular or not applicable | |
| Stipe is bare | |
| Ecology is saprotrophic | |
| Edibility is unknown | |
Taxonomy and naming
The fungus, originally collected from Muskingum County, Ohio, was named Peziza occidentalis by Lewis David de Schweinitz in 1832.[2] It was assigned its current name by Pier Andrea Saccardo in 1888.[3] Andrew Price Morgan renamed the species Geopyxis occidentalis in 1902 because of a perceived similarity with Geopyxis hesperidea,[4] but the name change was not adopted by subsequent authors. In 1928, Fred Jay Seaver overturned Saccardo's naming and applied the name Plectania to Sarcoscypha coccinea and other red cup fungi.[5] In later taxonomic revisions, Richard P. Korf reinstated the genus name Sarcoscypha.[6][7]
The specific epithet occidentalis, derived from the Latin word for "western", may refer to the distribution of the species in the western hemisphere. It is commonly known as the stalked scarlet cup[8] or the western scarlet cup.[9]
Phylogeny
| ||||||||||||||||||||||||||||||||||||||||||
| Phylogeny and relationships of S. occidentalis and related species based on ITS sequences and morphological characteristics.[10] |
The phylogenetic relationships in the genus Sarcoscypha were analyzed by Francis Harrington in the late 1990s.[10][11] The cladistic analysis combined comparison of sequences from the internal transcribed spacer in the non-functional RNA with fifteen traditional morphological characters, such as spore features, fruit body shape, and degree of hair curliness. Based on this analysis, S. occidentalis is part of a clade of evolutionarily related taxa that includes the species S. dudleyi, S. emarginata, S. hosoyae, S. korfiana and S. mesocyatha.[10] All of these species contain large oil droplets in their spores, in contrast to the other major clade of Sarcoscypha (containing the type species S. coccinea), characterized by having smaller, more numerous droplets.[11] The species most closely related to S. occidentalis is S. mesocyatha, known only from Hawaii.[12]
Description

Depending on their age, the fruit bodies of S. occidentalis may range in shape from deep cups to saucers to discs in maturity, and they can reach diameters up to 2 cm (0.8 in). In young specimens, the edges of the cup are curled inwards, and crenulate (with small rounded scallops); the cup edges in older specimens become laciniate (with jagged edges cut into irregular segments). The cups rest atop a stem that is small to medium-sized, up to 30 mm (1.2 in) long and 1.5–2 mm thick,[13] and attached centrally or to the side to the underside of the cup. The base of the stem may be covered with translucent "hairs".[14] The fertile spore-bearing inner surface of the cups, the hymenium, is bright red but fades to yellow or orange when dry. It is smooth or becomes so with time.[15] The fruit bodies are fleshy to rubbery when fresh, but become leathery when dry.[13] The flesh is thin and has no distinctive odor or taste. S. occidentalis is "of no culinary value".[8] A Jamaican variety has been named (as Plectania occidentalis var. jamaicensis); it has a pinker hymenium.[16]
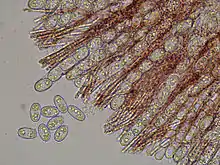
Exipulum is a term used to refer to the tissue or tissues containing the hymenium of an ascomycete fruit body. The ectal excipulum (outer tissue layer) is thin (20–30 µm thickness), made of a tissue type known as texura porrecta, consisting of more or less parallel hyphae all in one direction, with wide lumina and non-thickened walls. The medullary exipulum (middle tissue layer) is thick (200–600 µm) and made of textura intricata, a tissue layer made of irregularly interwoven hyphae with distinct spaces between the hyphae. The asci (filamentous structures in which the ascospores develop) are cylindrical with gradually tapering bases, eight-spored, and measure 240–280 by 12–15 µm. The ascospores have ellipsoidal to roughly cylindrical shapes, usually with blunt ends, and measure 19–22 by 10–12 µm. They have smooth surfaces and usually contain two large oil drops. The paraphyses (sterile, filamentous hyphae present in the hymenium) are cylindrical, 2–3 µm thick, barely enlarged at their apices, straight, and mostly unbranched above. They may sometimes anastomose, but do not form a conspicuous network.[15] The paraphyses contain numerous red granules.[17]
Anamorph form
Anamorphic or imperfect fungi are those that seem to lack a sexual stage in their life cycle, and typically reproduce by the process of mitosis in structures called conidia. In some cases, the sexual stage—or teleomorph stage—is later identified, and a teleomorph-anamorph relationship is established between the species. The International Code of Botanical Nomenclature permits the recognition of two (or more) names for one and the same organisms, one based on the teleomorph, the other(s) restricted to the anamorph. The anamorphic state of S. occidentalis is Molliardiomyces occidentalis, described by John W. Paden. This form produces smooth, colorless conidiophores (specialized stalks that bear conidia) measuring 20–230 by 2–3.2 µm. The conidia are roughly spherical to ovoid, smooth, translucent (hyaline), and 4.6–7.0 by 3.0–3.8 µm.[18]
Similar species


Sarcoscypha occidentalis is frequently confused with S. coccinea, but is distinguished macroscopically from this species by its smaller fruit bodies, smaller spores, and less hairy exterior. The two also differ in seasonal and geographic distribution: S. coccinea fruits earlier in the year, and is distributed in eastern North America, in the midwest, in the valleys between the Pacific coast and the Sierras and Cascades, as well as Europe, Africa, Australia, and India.[15] Another eastern North American species, S. austriaca, has scarlet fruit bodies up to 6 cm (2.4 in) wide, and fruits in the early spring.[19]
S. occidentalis may also be mistaken for Microstoma floccosum, which occurs in the same habitat. M. floccosum, however, has taller cups and is covered with stiff white hairs.[20] Another cup-fungus, Scutellinia scutellata, is disc-shaped without a stem, and is fringed with black hairs around its rim.[8]
Ecology, habitat and distribution
As a saprobic fungus, Sarcoscypha occidentalis is part of a community of fungi that play an important role in the forest ecosystem by breaking down the complex insoluble molecules cellulose and lignin of wood and leaf litter into smaller oligosaccharides that may be used by a variety of microbes.[21] Fruit bodies of S. coccinea may grow either solitarily, scattered or grouped together on sticks, twigs, and fragments of dead wood,[15] usually somewhat decomposed and partially buried in the top 5 cm (2.0 in) of soil and forest litter. It prefers soil that is moist and shaded and has a high content of humus. Like all Sarcoscypha species, it prefers the wood of angiosperms, such as oak, maple, and basswood;[13] one field guide notes a preference for shagbark hickory.[17] In the United States, it fruits from late spring to early autumn, which contrasts it with S. coccinea, found usually in the spring.[13]
The fungus is found in North America east of the Rocky Mountains, and at higher elevations in Central America and the Caribbean.[15] It has also been collected in Japan[22] and Taiwan.[23]
References
- "Species synonymy: Sarcoscypha occidentalis (Schwein.) Sacc". Species Fungorum. CAB International. Retrieved 2010-04-02.
- von Schweinitz LD. (1832). "Synopsis fungorum in America boreali media degentium". Transactions of the American Philosophical Society (in Latin). 4 (2): 141–316. doi:10.2307/1004834. JSTOR 1004834.
- Saccardo PA. (1888). "Discomyceteae et Phymatosphaeriaceae". Sylloge Fungorum (in Latin). 8: 154.
- Morgan AP. (1902). "The Discomycetes of the Miami Valley, Ohio". The Journal of Mycology. 8 (4): 179–92. doi:10.2307/3752419. JSTOR 3752419.
- Seaver FJ. (1928). The North American cup-fungi (Operculates). New York, New York: Self published. p. 193.
- Korf RP. (1953). "The new rules of typification as they affect Sarcoscypha and Velutaria". Mycologia. 45 (2): 296–301. doi:10.1080/00275514.1953.12024269. JSTOR 4547696.
- Korf RP. (1972). "A synoptic key to the genera of the Pezizales". Mycologia. 64 (5): 937–94. doi:10.2307/3758070. JSTOR 3758070.
- Roody WC. (2003). Mushrooms of West Virginia and the Central Appalachians. Lexington, Kentucky: University Press of Kentucky. p. 470. ISBN 0-8131-9039-8.
- McKnight VB; McKnight KH. (1987). A Field Guide to Mushrooms: North America. Boston, Massachusetts: Houghton Mifflin. p. 208. ISBN 0-395-91090-0.
- Harrington FA. (1998). "Relationships among Sarcoscypha species: evidence from molecular and morphological characters". Mycologia. 90 (2): 235–43. doi:10.2307/3761299. JSTOR 3761299.
- Harrington FA; Potter D. (1997). "Phylogenetic relationships within Sarcoscypha based upon nucleotide sequences of the internal transcribed spacer of nuclear ribosomal DNA". Mycologia. 89 (2): 258–67. doi:10.2307/3761080. JSTOR 3761080.
- Hemmes DE; Desjardin D. (2002). Mushrooms of Hawai'i: An Identification Guide. Berkeley, California: Ten Speed Press. p. 141. ISBN 1-58008-339-0.
- Harrington FA. (1990). "Sarcoscypha in North America (Pezizales, Sarcoscyphaceae)". Mycotaxon. 38: 417–58.
- Kanouse BC. (1948). "The genus Plectania and its segregates in North America". Mycologia. 40 (4): 482–97. doi:10.2307/3755155. JSTOR 3755155.
- Denison WC. (1972). "Central American Pezizales. IV. The genera Sarcoscypha, Pithya, and Nanoscypha". Mycologia. 64 (3): 609–23. doi:10.2307/3757876. JSTOR 3757876.
- Dennis RWG. (1954). "Operculate Discomycetes from Trinidad and Tobago". Kew Bulletin. 9 (3): 417–21. doi:10.2307/4108810. JSTOR 4108810.
- Healy RA; Huffman DR; Tiffany LH; Knaphaus G. (2008). Mushrooms and Other Fungi of the Midcontinental United States. Bur Oak Guide. Iowa City, Iowa: University of Iowa Press. p. 291. ISBN 978-1-58729-627-7.
- Paden JW. (1984). "A new genus of Hyphomycetes with teleomorphs in the Sarcoscyphaceae (Pezizales, Sarcoscyphineae)". Canadian Journal of Botany. 62 (2): 211–8. doi:10.1139/b84-035.
- Miller HR; Miller OK. (2006). North American Mushrooms: a Field Guide to Edible and Inedible Fungi. Guilford, Connecticut: Falcon Guide. p. 536. ISBN 0-7627-3109-5.
- Seaver FJ. (1904). "The Discomycetes of eastern Iowa". Bulletin from the Laboratories of Natural History of the State University of Iowa: 60. ISBN 9780608416786.
- Edwards IP; Upchurch RA; Zak DR. (2008). "Isolation of fungal cellobiohydrolase I genes from sporocarps and forest soils by PCR". Applied and Environmental Microbiology. 74 (11): 3481–89. Bibcode:2008ApEnM..74.3481E. doi:10.1128/AEM.02893-07. PMC 2423022. PMID 18408067.
- Nagao H; Fukiharu T. (2002). "Fungal flora in Chiba Pref., central Japan (III) ascomycetes: Plectomycetes and Discomycetes". Journal of the Natural History Museum and Institute, Chiba (in Japanese) (5): 111–32. ISSN 1340-2684.
- Lious S-C; Chen Z-C. (1977). "Notes on Taiwan Discomycetes. Part I. Pezizales and Helotiales" (PDF). Taiwania. 22 (1): 29–43. ISSN 0372-333X.