Silicon isotope biogeochemistry
Silicon isotope biogeochemistry is the study of environmental processes using the relative abundance of Si isotopes. As the relative abundance of Si stable isotopes varies among different natural materials,[2] the differences in abundance can be used to trace the source of Si, and to study biological, geological, and chemical processes.[1] The study of stable isotope biogeochemistry of Si aims to quantify the different Si fluxes in the global biogeochemical silicon cycle, to understand the role of biogenic silica within the global Si cycle, and to investigate the applications and limitations of the sedimentary Si record as an environmental and palaeoceanographic proxy.[1]
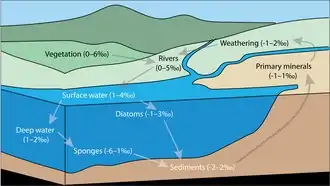
Background
Silicon in nature is typically bonded to oxygen, in a tetravalent oxidation state. The major forms of solid Si are silicate minerals and amorphous silica, whereas in aqueous solutions the dominant forms are orthosilicic acid and its dissociated species.[3] There are three stable isotopes of Si, associated with the following mean natural abundances: 28Si– 92.23%, 29Si– 4.67%, and 30Si– 3.10%.[2] The isotopic composition of Si is often formulated by the delta notation, as the following:

The reference material (standard) for defining the δ30Si of a sample is the National Bureau of Standards (NBS) 28 Sand Quartz, which has been certified and distributed by the National Institute of Standards and Technology (NIST), and is also named NIST RM 8546.[3] Currently, there are four main analytical methods for the measurement of Si isotopes: Gas Source Isotope-Ratio Mass Spectrometry (GC-IRMS), Secondary Ion Mass Spectrometry (SIMS), Multi-Collector Inductively Coupled Plasma Mass Spectrometry (MC–IPC–MS), and Laser Ablation MC–ICP–MS.[3]
Si isotopes in the Si biogeochemical cycle
Primary minerals and weathering
Primary minerals are the minerals that crystalize during the formation of Earth's crust, and their typical δ30Si isotopic value is in the range of −0.9‰ – +1.4‰.[1] Earth's crust is constantly undergoing weathering processes, which dissolve Si and produce secondary Si minerals simultaneously. The formation of secondary Si discriminates against the heavy Si isotope (30Si), creating minerals with relatively low δ30Si isotopic values (−3‰ – +2.5‰, mean: −1.1‰).[4] It has been suggested that this isotopic fractionation is controlled by the kinetic isotope effect of Si adsorption to Aluminum hydroxides, which takes place in early stages of weathering.[5] As a result of incorporation of lighter Si isotopes into secondary minerals, the remaining dissolved Si will be relative enriched in the heavy Si isotope (30Si), and associated with relatively high δ30Si isotopic values (−1‰ – +2‰, mean: +0.8‰).[4] The dissolved Si is often transported by rivers to the oceans.
Terrestrial vegetation
Silicon uptake by plants typically discriminates against the light Si isotope, forming 30Si-enriched plants (δ30Si of 0–6‰).[1] The reason for this relatively large isotopic fractionation remains unclear, mainly because the mechanisms of Si uptake by plants are yet to be understood. Silicon in plants can be found in the xylem, which is associated with exceptionally high δ30Si values.[6] Phytoliths, microscopic structures of silica in plant tissues, have relatively lower δ30Si values.[6] For example, it was reported that the mean δ30Si of phytoliths in various wheat organs were -1.4–2.1‰,[7] which is lower than the typical range for vegetation (δ30Si of 0–6‰). Phytoliths are relatively soluble, and as plants decay they contribute to the terrestrial dissolved Si budget.[1]
Biomineralization in aquatic environments
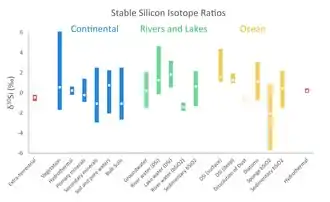
In aquatic environments (rivers, lakes and ocean), dissolved Si is utilized by diatoms, dictyochales, radiolarians and sponges to produce solid bSiO2 structures. The biomineralized silica has an amorphous structure and therefore its properties may vary among the different organisms.[8] Biomineralization by diatoms induces the largest Si flux within the ocean, and thus it has a crucial role in the global Si cycle.[9] During Si uptake by diatoms, there is an isotopic discrimination against the heavy isotope, forming 30Si-depleted biogenic silica minerals.[10] As a result, the remaining dissolved Si in the surrounding water is 30Si-enriched. Since diatoms rely on sunlight for photosynthesis, they inhabit in surface waters, and thus the surface water of the ocean are typically 30Si-enriched.[1] Although there is less available data on the isotopic fractionation during biomineralization by radiolarians, it has been suggested that radiolarians also discriminate against the heavy isotope (30Si), and that the magnitude of isotopic fractionation is of a similar range as biomineralization by diatoms.[11] Sponges also show an isotopic preference for 28Si over 30Si, but the magnitude of their isotopic fractionation is often larger[1] (For quantitative comparation, see Figure 2).
Hydrothermal vents
Hydrothermal vents contribute dissolved Si to the ocean Si reservoir. Currently, it is challenging to determine the magnitude of hydrothermal Si fluxes, due to lack of data on the δ30Si values associated with this flux.[1] There are only two published data points of the δ30Si value of hydrothermal vents (−0.4‰ and −0.2‰).[12]
Diagenesis
The δ30Si value of sediment porewater may be affected by post-depositional (diagenetic) precipitation or dissolution of Si. It is important to understand the extent and isotopic fractionations of these processes, as they alter the δ30Si values of the originally deposited sediments, and determine the δ30Si preserved in the rock record.[1] Generally, precipitation of Si prefers the light isotope (28Si) and leads to 30Si-enriched dissolved Si in the hosting solution.[13] The isotopic effect of Si dissolution in porewater is yet to be clear, as some studies report a preference for 28Si during dissolution,[14] while other studies document that isotopic fractionation was not expressed during dissolution of sediments.[15]
Paleoceanography proxies
The silicic acid leakage hypothesis
The silicic acid leakage hypothesis (SALH) is a suggested mechanism that aims to explain the atmospheric CO2 variations between glacial and interglacial periods.[16] This hypothesis proposes that during glacial periods, as a result of enhanced dust deposition in the southern ocean, diatoms consume less Si relative to nitrogen. The decrease in the Si:N uptake ratios leads to Si excess in the southern ocean, which leaks to lower latitudes of the ocean that are dominated by coccolithophores. As the Si concentrations rise, the diatom population may outcompete the coccolithophores, reducing the CaCO3 precipitation and altering ocean alkalinity and the carbonate pump.[17] These changes would induce a new ocean-atmosphere steady state with lower atmospheric CO2 concentrations, consistent with the draw down of CO2 observed in the last glacial period.[16] The δ30Si and δ15N isotopic values archived in the southern ocean diatom sediments has been used to examine this hypothesis,[18] as the dynamics of Si and N supply and utilization during the last deglaciation could be interpreted from this record. In alignment with the silicic acid leakage hypothesis, these isotopic archives suggest that Si utilization in the southern ocean increased during the deglaciation.[18]
Si isotope palaeothermometry
There have been attempts to reconstruct ocean paleotemperatures by chert Si isotopic record, which proposed that the Archean seawater temperatures were significantly higher than modern (~70 °C).[19] However, subsequent studies question this palaeothermometry method and offer alternative explanation for the δ30Si values of Archean rocks.[3] These signals could result from diagenetic alteration processes that overprint the original δ30Si values,[20] or reflect that Archean cherts were composed of different Si sources. It is plausible that in during the Archean the dominant sources of Si sediments were weathering, erosion, silicification of clastic sediments or hydrothermal activity,[21] in contrast to the vast SiO2 biomineralization in the modern ocean.
Paleo Si concentrations
According to empirical calibrations, the difference in δ30Si (denoted as Δ30Si) between sponges and their hosting water is correlated with the Si concentration of the hosting solution.[22] Therefore, it has been suggested that the Si concentrations in bottom waters of ancient oceans can be interpreted from the δ30Si of coexisting sponge spicules, which are preserved in the rock record.[22] It has been proposed that this relation is determined by the growth rate and the Si uptake kinetics of sponges,[22] but the current understanding of sponge biomineralization pathways is limited.[1] Although the mechanism behind this relation is yet to be clear, it appears consistent among various laboratory experiments, modern environments, and core top sediments.[1] However, there is also evidence that the δ30Si of carnivorous sponges may differ significantly from the expected correlation.[23]
See also
References
- Sutton, Jill N.; André, Luc; Cardinal, Damien; Conley, Daniel J.; de Souza, Gregory F.; Dean, Jonathan; Dodd, Justin; Ehlert, Claudia; Ellwood, Michael J.; Frings, Patrick J.; Grasse, Patricia (2018). "A Review of the Stable Isotope Bio-geochemistry of the Global Silicon Cycle and Its Associated Trace Elements". Frontiers in Earth Science. 5: 112. Bibcode:2018FrEaS...5..112S. doi:10.3389/feart.2017.00112. ISSN 2296-6463.
- De Biévre, P.; Taylor, P. D. P. (1993-02-04). "Table of the isotopic compositions of the elements". International Journal of Mass Spectrometry and Ion Processes. 123 (2): 149–166. Bibcode:1993IJMSI.123..149D. doi:10.1016/0168-1176(93)87009-H. ISSN 0168-1176.
- Poitrasson, Franck (2017-01-01). "Silicon Isotope Geochemistry". Reviews in Mineralogy and Geochemistry. 82 (1): 289–344. Bibcode:2017RvMG...82..289P. doi:10.2138/rmg.2017.82.8. ISSN 1529-6466.
- Frings, Patrick J.; Clymans, Wim; Fontorbe, Guillaume; De La Rocha, Christina L.; Conley, Daniel J. (2016-05-01). "The continental Si cycle and its impact on the ocean Si isotope budget". Chemical Geology. 425: 12–36. Bibcode:2016ChGeo.425...12F. doi:10.1016/j.chemgeo.2016.01.020. ISSN 0009-2541. S2CID 52043719.
- Oelze, Marcus; von Blanckenburg, Friedhelm; Hoellen, Daniel; Dietzel, Martin; Bouchez, Julien (2014-07-25). "Si stable isotope fractionation during adsorption and the competition between kinetic and equilibrium isotope fractionation: Implications for weathering systems". Chemical Geology. 380: 161–171. Bibcode:2014ChGeo.380..161O. doi:10.1016/j.chemgeo.2014.04.027. ISSN 0009-2541. S2CID 95924835.
- Opfergelt, S.; Cardinal, D.; Henriet, C.; André, L.; Delvaux, B. (2006-01-01). "Silicon isotope fractionation between plant parts in banana: In situ vs. in vitro". Journal of Geochemical Exploration. Extended Abstracts presented at the 7th Symp. on the Geochemistry of the Earth's Surface (GES-7). 88 (1): 224–227. Bibcode:2006JCExp..88..224O. doi:10.1016/j.gexplo.2005.08.044. ISSN 0375-6742.
- Hodson, Martin J.; Parker, Adrian G.; Leng, Melanie J.; Sloane, Hilary J. (May 2008). "Silicon, oxygen and carbon isotope composition of wheat (Triticum aestivum L.) phytoliths: implications for palaeoecology and archaeology". Journal of Quaternary Science. 23 (4): 331–339. Bibcode:2008JQS....23..331H. doi:10.1002/jqs.1176. S2CID 85096150.
- Perry, Carole C.; Belton, David; Shafran, Kirill (2003), Müller, Werner E. G. (ed.), "Studies of Biosilicas; Structural Aspects, ChemicalPrinciples,Model Studies and the Future", Silicon Biomineralization: Biology — Biochemistry — Molecular Biology — Biotechnology, Berlin, Heidelberg: Springer, vol. 33, pp. 269–299, doi:10.1007/978-3-642-55486-5_11, ISBN 978-3-642-55486-5, PMID 14518377, retrieved 2022-05-22
- Tréguer, Paul J.; De La Rocha, Christina L. (2013-01-03). "The World Ocean Silica Cycle". Annual Review of Marine Science. 5 (1): 477–501. doi:10.1146/annurev-marine-121211-172346. ISSN 1941-1405. PMID 22809182.
- Sutton, Jill N.; Varela, Diana E.; Brzezinski, Mark A.; Beucher, Charlotte P. (2013-03-01). "Species-dependent silicon isotope fractionation by marine diatoms". Geochimica et Cosmochimica Acta. 104: 300–309. Bibcode:2013GeCoA.104..300S. doi:10.1016/j.gca.2012.10.057. ISSN 0016-7037.
- Abelmann, Andrea; Gersonde, Rainer; Knorr, Gregor; Zhang, Xu; Chapligin, Bernhard; Maier, Edith; Esper, Oliver; Friedrichsen, Hans; Lohmann, Gerrit; Meyer, Hanno; Tiedemann, Ralf (2015-09-18). "The seasonal sea-ice zone in the glacial Southern Ocean as a carbon sink". Nature Communications. 6 (1): 8136. Bibcode:2015NatCo...6.8136A. doi:10.1038/ncomms9136. ISSN 2041-1723. PMC 4595604. PMID 26382319.
- De La Rocha, Christina L.; DePaolo, Donald J. (2000-08-18). "Isotopic Evidence for Variations in the Marine Calcium Cycle Over the Cenozoic". Science. 289 (5482): 1176–1178. Bibcode:2000Sci...289.1176D. doi:10.1126/science.289.5482.1176. ISSN 0036-8075. PMID 10947981.
- Geilert, Sonja; Vroon, Pieter Z.; Roerdink, Desiree L.; Van Cappellen, Philippe; van Bergen, Manfred J. (2014-10-01). "Silicon isotope fractionation during abiotic silica precipitation at low temperatures: Inferences from flow-through experiments". Geochimica et Cosmochimica Acta. 142: 95–114. Bibcode:2014GeCoA.142...95G. doi:10.1016/j.gca.2014.07.003. ISSN 0016-7037.
- Demarest, Mark S.; Brzezinski, Mark A.; Beucher, Charlotte P. (2009-10-01). "Fractionation of silicon isotopes during biogenic silica dissolution". Geochimica et Cosmochimica Acta. 73 (19): 5572–5583. Bibcode:2009GeCoA..73.5572D. doi:10.1016/j.gca.2009.06.019. ISSN 0016-7037.
- Wetzel, F.; de Souza, G.F.; Reynolds, B.C. (April 2014). "What controls silicon isotope fractionation during dissolution of diatom opal?". Geochimica et Cosmochimica Acta. 131: 128–137. Bibcode:2014GeCoA.131..128W. doi:10.1016/j.gca.2014.01.028.
- Matsumoto, Katsumi; Sarmiento, Jorge L.; Brzezinski, Mark A. (September 2002). "Silicic acid leakage from the Southern Ocean: A possible explanation for glacial atmospheric p CO 2: SOUTHERN OCEAN SILICIC ACID LEAKAGE". Global Biogeochemical Cycles. 16 (3): 5–1–5–23. doi:10.1029/2001GB001442. S2CID 10029685.
- Matsumoto, Katsumi; Sarmiento, Jorge L. (June 2008). "A corollary to the silicic acid leakage hypothesis: SALH COROLLARY". Paleoceanography. 23 (2): n/a. doi:10.1029/2007PA001515.
- Horn, Matthew G.; Beucher, Charlotte P.; Robinson, Rebecca S.; Brzezinski, Mark A. (2011-10-15). "Southern ocean nitrogen and silicon dynamics during the last deglaciation". Earth and Planetary Science Letters. 310 (3): 334–339. Bibcode:2011E&PSL.310..334H. doi:10.1016/j.epsl.2011.08.016. ISSN 0012-821X.
- Robert, François; Chaussidon, Marc (October 2006). "A palaeotemperature curve for the Precambrian oceans based on silicon isotopes in cherts". Nature. 443 (7114): 969–972. Bibcode:2006Natur.443..969R. doi:10.1038/nature05239. ISSN 1476-4687. PMID 17066030. S2CID 4417157.
- Stefurak, Elizabeth J. T.; Fischer, Woodward W.; Lowe, Donald R. (2015-02-01). "Texture-specific Si isotope variations in Barberton Greenstone Belt cherts record low temperature fractionations in early Archean seawater". Geochimica et Cosmochimica Acta. 150: 26–52. Bibcode:2015GeCoA.150...26S. doi:10.1016/j.gca.2014.11.014. ISSN 0016-7037.
- van den Boorn, Sander H.J.M.; van Bergen, Manfred J.; Nijman, Wouter; Vroon, Pieter Z. (2007-10-01). "Dual role of seawater and hydrothermal fluids in Early Archean chert formation: Evidence from silicon isotopes". Geology. 35 (10): 939–942. Bibcode:2007Geo....35..939V. doi:10.1130/G24096A.1. hdl:1874/386738. ISSN 0091-7613. S2CID 129800000.
- Hendry, Katharine R.; Robinson, Laura F. (2012-03-15). "The relationship between silicon isotope fractionation in sponges and silicic acid concentration: Modern and core-top studies of biogenic opal". Geochimica et Cosmochimica Acta. 81: 1–12. Bibcode:2012GeCoA..81....1H. doi:10.1016/j.gca.2011.12.010. hdl:1912/5093. ISSN 0016-7037. S2CID 53119488.
- Hendry, K. R.; Swann, G. E. A.; Leng, M. J.; Sloane, H. J.; Goodwin, C.; Berman, J.; Maldonado, M. (2015-06-05). "Technical Note: Silica stable isotopes and silicification in a carnivorous sponge Asbestopluma sp". Biogeosciences. 12 (11): 3489–3498. Bibcode:2015BGeo...12.3489H. doi:10.5194/bg-12-3489-2015. ISSN 1726-4170. S2CID 53051498.