Y RNA
Y RNAs are small non-coding RNAs. They are components of the Ro60 ribonucleoprotein particle[1] which is a target of autoimmune antibodies in patients with systemic lupus erythematosus.[2] They are also reported to be necessary for DNA replication through interactions with chromatin and initiation proteins.[3][4] However, mouse embryonic stem cells lacking Y RNAs are viable and have normal cell cycles.[5]
| Y RNA | |
|---|---|
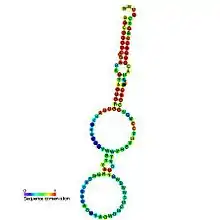 Predicted secondary structure and sequence conservation of Y_RNA | |
| Identifiers | |
| Symbol | Y_RNA |
| Alt. Symbols | Y1; Y3; Y4; Y5 |
| Rfam | RF00019 |
| Other data | |
| RNA type | Gene |
| Domain(s) | Eukaryota |
| SO | SO:0000405 |
| PDB structures | PDBe |
Structure
These small RNAs are predicted to fold into a conserved stem formed by the RNA's 3′ and 5′ ends and characterized by a single bulged cytosine, which are the known requirements for Ro binding.[6][7][8]
Function
Two functions have been described for Y RNAs in the literature: As a repressor of Ro60, and as an initiation factor for DNA replication. Mutant human Y RNAs lacking the conserved binding site for Ro60 protein still support DNA replication,[3] indicating that binding to Ro protein and promoting DNA replication are two separable functions of Y RNAs. Although Y RNA-derived small RNAs are similar in size to microRNAs, it has been shown that these Y RNA fragments are not involved in the microRNA pathway.[9]

Ro60 Inhibition
In its free state, Ro binds to a variety of misfolded RNAs including misfolded 5S rRNAs, and is thought to act as some sort of quality control mechanism.[10] Crystal structures of Ro complexed either with Y RNA or another RNA showed that Ro binds single-stranded 3′ ends of RNAs relatively nonspecifically, whereas Y RNA binds specifically at a second site that regulates access of other RNAs.[6] In Deinococcus, free Ro has also been shown to function in 23S rRNA maturation.[11] In Deinococcus, mutants lacking Y RNA are viable, and Y RNA appears to be unstable except when complexed with Ro.[11]
DNA replication initiation
Human Y RNAs are functionally required for DNA replication.[3] Biochemical fractionation and reconstitution experiments have established a functional requirement of human Y RNAs for chromosomal DNA replication in isolated vertebrate cell nuclei in vitro[3] and specific degradation of human Y RNAs inhibits DNA replication in vitro, and in intact cells in vivo.[3] Y RNA function is thought to be mediated via interactions with chromatin and initiation proteins (including the origin recognition complex)[4]
In human pathology
Y RNAs are overexpressed in some human tumours and are required for cell proliferation[12] and small, microRNA-sized breakdown products may be involved in autoimmunity and other pathological conditions.[13] Recent work has demonstrated that Y RNAs are modified at their 3′ end by the non-canonical poly(A) polymerase PAPD5, and the short oligo(A) tail added by PAPD5 is a marker for 3′ end processing by the ribonuclease PARN/EXOSC10 or for degradation by the exonuclease DIS3L.[14] Since PARN deficiency causes a severe form of the bone marrow disease Dyskeratosis Congenita as well as pulmonary fibrosis,[15][16] it is possible that defects in Y RNA processing contribute to the severe pathology observed in these patients.
Species distribution
Presumptive Y RNA and Ro protein homologs have been found in eukaryotes and bacteria.[7][17]
Humans
Humans appear to have four Y RNAs, named hY1, hY3, hY4 and hY5[17] and also a large number of pseudogenes.
C. elegans
Caenorhabditis elegans has one, named CeY RNA and a large number of sbRNAs that are postulated to also be Y RNA homologues.[18][19]
D. radiodurans
The radiation-resistant bacterium Deinococcus radiodurans encodes a homolog of Ro called rsr ("Ro sixty related"), and at least four small RNAs accumulate in Deinococcus under conditions where rsr expression is induced (UV irradiation); one of these RNAs appears to be a Y RNA homolog.[20] In Deinococcus radiodurans Rsr is tethered via Y RNA to the exoribonuclease PNPase and channels single-stranded RNA into the PNPase cavity. Rsr and Y RNA enhance degradation of structured RNAs by PNPase. This role could be conserved, as Rsr and ncRNAs called YrlA and YrlB (Y RNA like) also associate with PNPase in an evolutionary distant bacterium Salmonella Typhimurium.[21]
References
- Hall AE, Turnbull C, Dalmay T (April 2013). "Y RNAs: recent developments". Biomolecular Concepts. 4 (2): 103–110. doi:10.1515/bmc-2012-0050. PMID 25436569. S2CID 12575326.
- Lerner MR, Boyle JA, Hardin JA, Steitz JA (January 1981). "Two novel classes of small ribonucleoproteins detected by antibodies associated with lupus erythematosus". Science. 211 (4480): 400–402. Bibcode:1981Sci...211..400L. doi:10.1126/science.6164096. PMID 6164096.
- Christov CP, Gardiner TJ, Szüts D, Krude T (September 2006). "Functional requirement of noncoding Y RNAs for human chromosomal DNA replication". Molecular and Cellular Biology. 26 (18): 6993–7004. doi:10.1128/MCB.01060-06. PMC 1592862. PMID 16943439.
- Zhang AT, Langley AR, Christov CP, Kheir E, Shafee T, Gardiner TJ, Krude T (June 2011). "Dynamic interaction of Y RNAs with chromatin and initiation proteins during human DNA replication". Journal of Cell Science. 124 (Pt 12): 2058–2069. doi:10.1242/jcs.086561. PMC 3104036. PMID 21610089.
- Leng Y, Sim S, Magidson V, Wolin SL (July 2020). "Noncoding Y RNAs regulate the levels,subcellular distribution and protein interactions of their Ro60 autoantigen partner". Nucleic Acids Research. 48 (12): 6919–6930. doi:10.1093/nar/gkaa414. PMC 7337961. PMID 32469055.
- Stein AJ, Fuchs G, Fu C, Wolin SL, Reinisch KM (May 2005). "Structural insights into RNA quality control: the Ro autoantigen binds misfolded RNAs via its central cavity". Cell. 121 (4): 529–539. doi:10.1016/j.cell.2005.03.009. PMC 1769319. PMID 15907467.
- Teunissen SW, Kruithof MJ, Farris AD, Harley JB, Venrooij WJ, Pruijn GJ (January 2000). "Conserved features of Y RNAs: a comparison of experimentally derived secondary structures". Nucleic Acids Research. 28 (2): 610–619. doi:10.1093/nar/28.2.610. PMC 102524. PMID 10606662.
- Green CD, Long KS, Shi H, Wolin SL (July 1998). "Binding of the 60-kDa Ro autoantigen to Y RNAs: evidence for recognition in the major groove of a conserved helix". RNA. 4 (7): 750–765. doi:10.1017/S1355838298971667. PMC 1369656. PMID 9671049.
- Nicolas FE, Hall AE, Csorba T, Turnbull C, Dalmay T (April 2012). "Biogenesis of Y RNA-derived small RNAs is independent of the microRNA pathway". FEBS Letters. 586 (8): 1226–1230. doi:10.1016/j.febslet.2012.03.026. PMID 22575660. S2CID 7805517.
- Reinisch KM, Wolin SL (April 2007). "Emerging themes in non-coding RNA quality control". Current Opinion in Structural Biology. 17 (2): 209–214. doi:10.1016/j.sbi.2007.03.012. PMID 17395456.
- Chen X, Wurtmann EJ, Van Batavia J, Zybailov B, Washburn MP, Wolin SL (June 2007). "An ortholog of the Ro autoantigen functions in 23S rRNA maturation in D. radiodurans". Genes & Development. 21 (11): 1328–1339. doi:10.1101/gad.1548207. PMC 1877746. PMID 17510283.
- Christov CP, Trivier E, Krude T (March 2008). "Noncoding human Y RNAs are overexpressed in tumours and required for cell proliferation". British Journal of Cancer. 98 (5): 981–988. doi:10.1038/sj.bjc.6604254. PMC 2266855. PMID 18283318.
- Verhagen AP, Pruijn GJ (September 2011). "Are the Ro RNP-associated Y RNAs concealing microRNAs? Y RNA-derived miRNAs may be involved in autoimmunity". BioEssays. 33 (9): 674–682. doi:10.1002/bies.201100048. PMID 21735459. S2CID 26501077.
- Shukla S, Parker R (October 2017). "PARN Modulates Y RNA Stability and Its 3′-End Formation". Molecular and Cellular Biology. 37 (20). doi:10.1128/MCB.00264-17. PMC 5615183. PMID 28760775.
- Stuart BD, Choi J, Zaidi S, Xing C, Holohan B, Chen R, Choi M, Dharwadkar P, Torres F, Girod CE, Weissler J, Fitzgerald J, Kershaw C, Klesney-Tait J, Mageto Y, Shay JW, Ji W, Bilguvar K, Mane S, Lifton RP, Garcia CK (May 2015). "Exome sequencing links mutations in PARN and RTEL1 with familial pulmonary fibrosis and telomere shortening". Nature Genetics. 47 (5): 512–517. doi:10.1038/ng.3278. PMC 4414891. PMID 25848748.
- Tummala H, Walne A, Collopy L, Cardoso S, de la Fuente J, Lawson S, Powell J, Cooper N, Foster A, Mohammed S, Plagnol V, Vulliamy T, Dokal I (May 2015). "Poly(A)-specific ribonuclease deficiency impacts telomere biology and causes dyskeratosis congenita". The Journal of Clinical Investigation. 125 (5): 2151–2160. doi:10.1172/JCI78963. PMC 4463202. PMID 25893599.
- Perreault J, Perreault JP, Boire G (August 2007). "Ro-associated Y RNAs in metazoans: evolution and diversification". Molecular Biology and Evolution. 24 (8): 1678–1689. doi:10.1093/molbev/msm084. PMID 17470436.
- Van Horn DJ, Eisenberg D, O'Brien CA, Wolin SL (May 1995). "Caenorhabditis elegans embryos contain only one major species of Ro RNP". RNA. 1 (3): 293–303. PMC 1369082. PMID 7489501.
- Boria I, Gruber AR, Tanzer A, Bernhart SH, Lorenz R, Mueller MM, Hofacker IL, Stadler PF (April 2010). "Nematode sbRNAs: homologs of vertebrate Y RNAs". Journal of Molecular Evolution. 70 (4): 346–358. Bibcode:2010JMolE..70..346B. doi:10.1007/s00239-010-9332-4. PMID 20349053. S2CID 876486.
- Chen X, Quinn AM, Wolin SL (April 2000). "Ro ribonucleoproteins contribute to the resistance of Deinococcus radiodurans to ultraviolet irradiation". Genes & Development. 14 (7): 777–782. doi:10.1101/gad.14.7.777. PMC 316496. PMID 10766734.
- Chen X, Taylor DW, Fowler CC, Galan JE, Wang HW, Wolin SL (March 2013). "An RNA degradation machine sculpted by Ro autoantigen and noncoding RNA". Cell. 153 (1): 166–177. doi:10.1016/j.cell.2013.02.037. PMC 3646564. PMID 23540697.
External links
Types of nucleic acids | |||||||
|---|---|---|---|---|---|---|---|
| Constituents | |||||||
| Ribonucleic acids (coding, non-coding) |
| ||||||
| Deoxyribonucleic acids | |||||||
| Analogues | |||||||
| Cloning vectors | |||||||
| |||||||