Antimicrobial peptides
Antimicrobial peptides (AMPs), also called host defence peptides (HDPs) are part of the innate immune response found among all classes of life. Fundamental differences exist between prokaryotic and eukaryotic cells that may represent targets for antimicrobial peptides. These peptides are potent, broad spectrum antibiotics which demonstrate potential as novel therapeutic agents. Antimicrobial peptides have been demonstrated to kill Gram negative and Gram positive bacteria,[1] enveloped viruses, fungi and even transformed or cancerous cells.[2] Unlike the majority of conventional antibiotics it appears that antimicrobial peptides frequently destabilize biological membranes, can form transmembrane channels, and may also have the ability to enhance immunity by functioning as immunomodulators.
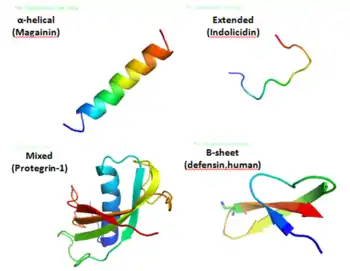
Structure

Antimicrobial peptides are a unique and diverse group of molecules, which are divided into subgroups on the basis of their amino acid composition and structure.[3] Antimicrobial peptides are generally between 12 and 50 amino acids. These peptides include two or more positively charged residues provided by arginine, lysine or, in acidic environments, histidine, and a large proportion (generally >50%) of hydrophobic residues.[4][5][6] The secondary structures of these molecules follow 4 themes, including i) α-helical, ii) β-stranded due to the presence of 2 or more disulfide bonds, iii) β-hairpin or loop due to the presence of a single disulfide bond and/or cyclization of the peptide chain, and iv) extended.[7] Many of these peptides are unstructured in free solution, and fold into their final configuration upon partitioning into biological membranes. It contains hydrophilic amino acid residues aligned along one side and hydrophobic amino acid residues aligned along the opposite side of a helical molecule.[3] This amphipathicity of the antimicrobial peptides allows them to partition into the membrane lipid bilayer. The ability to associate with membranes is a definitive feature of antimicrobial peptides,[8][9] although membrane permeabilization is not necessary. These peptides have a variety of antimicrobial activities ranging from membrane permeabilization to action on a range of cytoplasmic targets.
| Type | characteristic | AMPs |
|---|---|---|
| Anionic peptides | rich in glutamic and aspartic acids | Maximin H5 from amphibians, dermcidin from humans |
| Linear cationic α-helical peptides | lack in cysteine | Cecropins, andropin, moricin, ceratotoxin and melittin from insects, Magainin, dermaseptin, bombinin, brevinin-1, esculentins and buforin II from amphibians, CAP18 from rabbits, LL37 from humans |
| Cationic peptide enriched for specific amino acid | rich in proline, arginine, phenylalanine, glycine, tryptophan | abaecin and drosocin, apidaecin, diptericin, and attacin from insects, prophenin from pigs, indolicidin from cattle. |
| Anionic/cationic peptides forming disulfide bonds | contain 1~3 disulfide bond |
|
Activities

The modes of action by which antimicrobial peptides kill microbes are varied,[10] and may differ for different bacterial species.[11] Some antimicrobial peptides kill both bacteria and fungi, e.g., psoriasin kills E. coli and several filamentous fungi.[12] The cytoplasmic membrane is a frequent target, but peptides may also interfere with DNA and protein synthesis, protein folding, and cell wall synthesis.[10] The initial contact between the peptide and the target organism is electrostatic, as most bacterial surfaces are anionic, or hydrophobic, such as in the antimicrobial peptide Piscidin. Their amino acid composition, amphipathicity, cationic charge and size allow them to attach to and insert into membrane bilayers to form pores by ‘barrel-stave’, ‘carpet’ or ‘toroidal-pore’ mechanisms. Alternately, they may penetrate into the cell to bind intracellular molecules which are crucial to cell living.[13] Intracellular binding models includes inhibition of cell wall synthesis, alteration of the cytoplasmic membrane, activation of autolysin, inhibition of DNA, RNA, and protein synthesis, and inhibition of certain enzymes. However, in many cases, the exact mechanism of killing is not known. One emerging technique for the study of such mechanisms is dual polarisation interferometry.[14][15] In contrast to many conventional antibiotics these peptides appear to be bactericidal[2] instead of bacteriostatic. In general the antimicrobial activity of these peptides is determined by measuring the minimal inhibitory concentration (MIC), which is the lowest concentration of drug that inhibits bacterial growth.[16]
AMPs can possess multiple activities including anti-gram-positive bacterial, anti-gram-negative bacterial, anti-fungal, anti-viral, anti-parasitic, and anti cancer activities. A big AMP functional analysis indicates that among all AMP activities, amphipathicity and charge, two major properties of AMPs, best distinguish between AMPs with and without anti-gram-negative bacterial activities.[17] This implies that being AMPs with anti-gram-negative bacterial activities may prefer or even require strong amphipathicity and net positive charge.
Immunomodulation
In addition to killing bacteria directly they have been demonstrated to have a number of immunomodulatory functions that may be involved in the clearance of infection, including the ability to alter host gene expression, act as chemokines and/or induce chemokine production, inhibiting lipopolysaccharide induced pro-inflammatory cytokine production, promoting wound healing, and modulating the responses of dendritic cells and cells of the adaptive immune response. Animal models indicate that host defense peptides are crucial for both prevention and clearance of infection. It appears as though many peptides initially isolated as and termed "antimicrobial peptides" have been shown to have more significant alternative functions in vivo (e.g. hepcidin[18]). Dusquetide for example is an immunomodulator that acts through p62, a protein involved in toll like receptor based signalling of infection. The peptide is being examined in a Phase III clinical trial by Soligenix (SGNX) to ascertain if it can assist in repair of radiation-induced damage to oral mucosa arising during cancer radiotherapy of the head and neck.[19]
Mechanisms of action

Antimicrobial peptides generally have a net positive charge, allowing them to interact with the negatively charged molecules exposed on bacteria and cancer cell surfaces, such as phospholipid phosphatidylserine, O-glycosylated mucins, sialylated gangliosides, and heparin sulfates. The mechanism of action of these peptides varies widely but can be simplified into two categories: membranolytic and non-membranolytic antimicrobial peptides.[20] The disruption of membranes by membranolytic antimicrobial peptides can be described by four models:[20]
- toroidal model
- disordered toroidal-pore model
- carpet model
- barrel stave model
Although the specifics of each mechanism differ, all propose peptide-induced membrane rupture, allowing cytoplasmic leakage that ultimately leads to death.
Recent work has painted a more complex picture of antimicrobial peptide activity. The non-membranolytic antimicrobial peptides may also function as metabolic inhibitors, directly interacting with DNA, RNA, protein synthesis, and inhibitors of cell wall synthesis or septum formation. They are also known to cause ribosomal aggregation and delocalize membrane proteins.
Adding a further layer of complexity, many natural antimicrobial peptides possess weak bactericidal activity. Rather than directly inhibit bacterial growth, they are now known to act in concert with the host immune system through mechanisms including chemokine induction, histamine release, and angiogenesis modulation. These immunomodulatory effects have only recently begun to receive attention.
Several methods have been used to determine the mechanisms of antimicrobial peptide activity.[11][13] In particular, solid-state NMR studies have provided an atomic-level resolution explanation of membrane disruption by antimicrobial peptides.[21][22] In more recent years, X-ray crystallography has been used to delineate in atomic detail how the family of plant defensins rupture membranes by identifying key phospholipids in the cell membranes of the pathogen.[23][24] Human defensins have been thought to act through a similar mechanism, targeting cell membrane lipids as part of their function. In fact human beta-defensin 2 have now been shown to kill the pathogenic fungi Candida albicans through interactions with specific phospholipids.[25] From the computational point of view, the molecular dynamics simulations can shed light in the molecular mechanism and the specific peptide interactions with lipids, ions and solvent.[26]
| Methods | Applications |
|---|---|
| Microscopy | to visualize the effects of antimicrobial peptides on microbial cells |
| Atomic emission spectroscopy | to detect loss of intracellular potassium (an indication that bacterial membrane integrity has been compromised) |
| Fluorescent dyes | to measure ability of antimicrobial peptides to permeabilize membrane vesicles |
| Ion channel formation | to assess the formation and stability of an antimicrobial-peptide-induced pore |
| Circular dichroism and orientated circular dichroism | to measure the orientation and secondary structure of an antimicrobial peptide bound to a lipid bilayer |
| Dual polarization interferometry | to measure the different mechanisms of antimicrobial peptides |
| Solid-state NMR spectroscopy | to measure the secondary structure, orientation and penetration of antimicrobial peptides into lipid bilayers in the biologically relevant liquid-crystalline state |
| Neutron and X-ray diffraction | to measure the diffraction patterns of peptide-induced pores within membranes in oriented multilayers or liquids |
| Molecular dynamics simulations | to study the molecular behaviour and search for specific peptide-lipid interactions |
| Mass spectrometry | to measure the proteomic response of microorganisms to antimicrobial peptides |
Therapeutic research and use
Antimicrobial peptides have been used as therapeutic agents; their use is generally limited to intravenous administration or topical applications due to their short half-lives. As of January 2018 the following antimicrobial peptides were in clinical use:[27]
- Bacitracin for pneumonia, topical
- Boceprevir, Hepatitis C (oral, cyclic peptide)
- Dalbavancin, bacterial infections, IV
- Daptomycin, bacterial infections, IV
- Enfuvirtide, HIV, subcutaneous injection
- Oritavancin, bacterial infections, IV
- Teicoplanin, bacterial infections, IV
- Telaprevir, Hepatitis C, oral cyclic peptide
- Telavancin, bacterial infection, IV
- Vancomycin, bacterial infection, IV.
- Guavanin 2, bacterial infection against Gram-positive and Gram-negative also.
Activity beyond antibacterial functions
AMPs have been observed having functions other than bacterial and fungal killing. These activities include antiviral effects, but also roles in host defence such as anticancer functions and roles in neurology.[28] This has led to a movement for re-branding AMPs as "Host-defence peptides" to encompass the broad scope of activities AMPs can have.[29]
Anticancer properties
Some cecropins (e.g. cecropin A, and cecropin B) have anticancer properties and are called anticancer peptides (ACPs).[30]: 3 Hybrid ACPs based on Cecropin A have been studied for anticancer properties.[30]: 7.1 The fruit fly Defensin prevents tumour growth, suspected to bind to tumour cells owing to cell membrane modifications common to most cancer cells, such as phosphatidylserine exposure.[31]
Antibiofilm properties
Cecropin A can destroy planktonic and sessile biofilm-forming uropathogenic E. coli (UPEC) cells, either alone or when combined with the antibiotic nalidixic acid, synergistically clearing infection in vivo (in the insect host Galleria mellonella) without off-target cytotoxicity. The multi-target mechanism of action involves outer membrane permeabilization followed by biofilm disruption triggered by the inhibition of efflux pump activity and interactions with extracellular and intracellular nucleic acids.[32]
Selectivity
In the competition of bacterial cells and host cells with the antimicrobial peptides, antimicrobial peptides will preferentially interact with the bacterial cell to the mammalian cells, which enables them to kill microorganisms without being significantly toxic to mammalian cells.[39] Selectivity is a very important feature of the antimicrobial peptides and it can guarantee their function as antibiotics in host defense systems.
With regard to cancer cells, they themselves also secrete human antimicrobial peptides including defensin, and in some cases, they are reported to be more resistant than the surrounding normal cells. Therefore, we cannot conclude that selectivity is always high against cancer cells.[40][41]
Factors
There are some factors that are closely related to the selectivity property of antimicrobial peptides, among which the cationic property contributes most. Since the surface of the bacterial membranes is more negatively charged than mammalian cells, antimicrobial peptides will show different affinities towards the bacterial membranes and mammalian cell membranes.[42]
In addition, there are also other factors that will affect the selectivity. It's well known that cholesterol is normally widely distributed in the mammalian cell membranes as a membrane stabilizing agents but absent in bacterial cell membranes; and the presence of these cholesterols will also generally reduce the activities of the antimicrobial peptides, due either to stabilization of the lipid bilayer or to interactions between cholesterol and the peptide. So the cholesterol in mammalian cells will protect the cells from attack by the antimicrobial peptides.[43]
Besides, the transmembrane potential is well known to affect peptide-lipid interactions.[44] There's an inside-negative transmembrane potential existing from the outer leaflet to the inner leaflet of the cell membranes and this inside-negative transmembrane potential will facilitate membrane permeabilization probably by facilitating the insertion of positively charged peptides into membranes. By comparison, the transmembrane potential of bacterial cells is more negative than that of normal mammalian cells, so bacterial membrane will be prone to be attacked by the positively charged antimicrobial peptides.
Similarly, it is also believed that increasing ionic strength,[43] which in general reduces the activity of most antimicrobial peptides, contributes partially to the selectivity of the antimicrobial peptides by weakening the electrostatic interactions required for the initial interaction.

Mechanism
The cell membranes of bacteria are rich in acidic phospholipids, such as phosphatidylglycerol and cardiolipin.[39][45] These phospholipid headgroups are heavily negatively charged. Therefore, the outmost leaflets of the bilayer which is exposed to the outside of the bacterial membranes are more attractive to the attack of the positively charged antimicrobial peptides. So the interaction between the positive charges of antimicrobial peptides and the negatively charged bacterial membranes is mainly the electrostatic interactions, which is the major driving force for cellular association. In addition, since antimicrobial peptides form structures with a positively charged face as well as a hydrophobic face, there are also some hydrophobic interactions between the hydrophobic regions of the antimicrobial peptides and the zwitterionic phospholipids (electrically neutral) surface of the bacterial membranes, which act only as a minor effect in this case.
In contrast, the outer part of the membranes of plants and mammals is mainly composed of lipids without any net charges since most of the lipids with negatively charged headgroups are principally sequestered into the inner leaflet of the plasma membranes.[42] Thus in the case of mammalian cells, the outer surfaces of the membranes are usually made of zwitterionic phosphatidylcholine and sphingomyelin, even though a small portion of the membrane's outer surfaces contain some negatively charged gangliosides. Therefore, the hydrophobic interaction between the hydrophobic face of amphipathic antimicrobial peptides and the zwitterionic phospholipids on the cell surface of mammalian cell membranes plays a major role in the formation of peptide-cell binding.[46] However, the hydrophobic interaction is relatively weak when compared to the electrostatic interaction, thus, the antimicrobial peptides will preferentially interact with bacterial membranes.
Dual polarisation interferometry has been used in vitro to study and quantify the association to headgroup, insertion into the bilayer, pore formation and eventual disruption of the membrane.[47][48]
Control
A lot of effort has been put into controlling cell selectivity. For example, attempts have been made to modify and optimize the physicochemical parameters of the peptides to control the selectivities, including net charge, helicity, hydrophobicity per residue (H), hydrophobic moment (μ) and the angle subtended by the positively charged polar helix face (Φ).[44] Other mechanisms like the introduction of D-amino acids and fluorinated amino acids in the hydrophobic phase are believed to break the secondary structure and thus reduce hydrophobic interaction with mammalian cells. It has also been found that Pro→Nlys substitution in Pro-containing β-turn antimicrobial peptides was a promising strategy for the design of new small bacterial cell-selective antimicrobial peptides with intracellular mechanisms of action.[49] It has been suggested that direct attachment of magainin to the substrate surface decreased nonspecific cell binding and led to improved detection limit for bacterial cells such as Salmonella and E. coli.[50]
Bacterial resistance
Bacteria use various resistance strategies to avoid antimicrobial peptide killing.[13] Some microorganisms alter net surface charges. Staphylococcus aureus transports D-alanine from the cytoplasm to the surface teichoic acid which reduces the net negative charge by introducing basic amino groups.[51] S. aureus also modifies its anionic membranes via MprF with L-lysine, increasing the positive net charge.[51] The interaction of antimicrobial peptides with membrane targets can be limited by capsule polysaccharide of Klebsiella pneumoniae.[52] Alterations occur in Lipid A. Salmonella species reduce the fluidity of their outer membrane by increasing hydrophobic interactions between an increased number of Lipid A acyl tails by adding myristate to Lipid A with 2-hydroxymyristate and forming hepta-acylated Lipid A by adding palmitate. The increased hydrophobic moment is thought to retard or abolish antimicrobial peptide insertion and pore formation. The residues undergo alteration in membrane proteins. In some Gram-negative bacteria, alteration in the production of outer membrane proteins correlates with resistance to killing by antimicrobial peptides.[53] Non-typeable Hemophilus influenzae transports AMPs into the interior of the cell, where they are degraded. Furthermore, H. influenzae remodels its membranes to make it appear as if the bacterium has already been successfully attacked by AMPs, protecting it from being attacked by more AMPs.[54] ATP-binding cassette transporters import antimicrobial peptides and the resistance-nodulation cell-division efflux pump exports antimicrobial peptides.[55] Both transporters have been associated with antimicrobial peptide resistance. Bacteria produce proteolytic enzymes, which may degrade antimicrobial peptides leading to their resistance.[56] Outer membrane vesicles produced by Gram-negative bacteria bind the antimicrobial peptides and sequester them away from the cells, thereby protecting the cells.[57] The outer membrane vesicles are also known to contain various proteases, peptidases and other lytic enzymes, which may have a role in degrading the extracellular peptide and nucleic acid molecules, which if allowed to reach to the bacterial cells may be dangerous for the cells. Cyclic-di-GMP signaling had also been involved in the regulation of antimicrobial peptide resistance in Pseudomonas aeruginosa[58]
While these examples show that resistance can evolve naturally, there is increasing concern that using pharmaceutical copies of antimicrobial peptides can make resistance happen more often and faster. In some cases, resistance to these peptides used as a pharmaceutical to treat medical problems can lead to resistance, not only to the medical application of the peptides, but to the physiological function of those peptides.[59][60]
The ‘Trojan Horse’ approach to solving this problem capitalizes on the innate need for iron by pathogens. “Smuggling” antimicrobials into the pathogen is accomplished by linking them to siderophores for transport. While simple in concept, it has taken many decades of work to accomplish the difficult hurdle of transporting antimicrobials across the cell membranes of pathogens. Lessons learned from the successes and failures of siderophore-conjugate drugs evaluated during the development of novel agents using the ‘Trojan horse’ approach have been reviewed.[61]
Examples

Antimicrobial peptides are produced by species across the tree of life, including:
- bacteria (e.g. bacteriocin, and many others)
- fungi (e.g. peptaibols, plectasin, and many others)
- cnidaria (e.g. hydramacin, aurelin)
- many from insects and arthropods (e.g. cecropin, attacin, melittin, mastoparan, drosomycin)[62]
- amphibia, frogs (magainin, dermaseptin, aurein, and others)[63][64]
- birds (e.g. avian defensins)[65]
- and mammals (e.g. cathelicidins, alpha- and beta-defensins, regIII peptides)
Research has increased in recent years to develop artificially-engineered mimics of antimicrobial peptides such as SNAPPs, in part due to the prohibitive cost of producing naturally-derived AMPs.[66] An example of this is the facially cationic peptide C18G, which was designed from the C-terminal domain of human platelet factor IV.[67] Currently, the most widely used antimicrobial peptide is nisin; being the only FDA approved antimicrobial peptide, it is commonly used as an artificial preservative.[68]
Bioinformatics
Several bioinformatic databases exist to catalogue antimicrobial peptides. The APD (the Antimicrobial Peptide Database) is the original and model database for antimicrobial peptides (https://aps.unmc.edu). Based on the APD, other databases have also been built, including ADAM (A Database of Anti-Microbial peptides),[69] BioPD (Biologically active Peptide Database), CAMP (Collection of sequences and structures of antimicrobial peptides),[70] DBAASP (Database of Antimicrobial Activity and Structure of Peptides), DRAMP(Data Repository of Antimicrobial Peptides),[71] and LAMP (Linking AMPs).
The Antimicrobial peptide databases may be divided into two categories on the basis of the source of peptides it contains, as specific databases and general databases. These databases have various tools for antimicrobial peptides analysis and prediction. For example, the APD has a widely used calculation interface. It also provides links to many other tools. CAMP contains AMP prediction, feature calculator, BLAST search, ClustalW, VAST, PRATT, Helical wheel etc. In addition, ADAM allows users to search or browse through AMP sequence-structure relationships. Antimicrobial peptides often encompass a wide range of categories such as antifungal, antibacterial, and antituberculosis peptides.
dbAMP:[72] Provides an online platform for exploring antimicrobial peptides with functional activities and physicochemical properties on transcriptome and proteome data. dbAMP is an online resource that addresses various topics such as annotations of antimicrobial peptides (AMPs) including sequence information, antimicrobial activities, post-translational modifications (PTMs), structural visualization, antimicrobial potency, target species with minimum inhibitory concentration (MIC), physicochemical properties, or AMP–protein interactions.
Tools such as PeptideRanker,[73] PeptideLocator,[74] and AntiMPmod[75][76] allow for the prediction of antimicrobial peptides while others have been developed to predict antifungal and anti-Tuberculosis activities.[77][78]
See also
- Aurein
- Bacteriocin
- Cathelicidin
- Copsin
- Diptericin
- Peripheral membrane proteins
- Virtual colony count
References
- Ageitos JM, Sánchez-Pérez A, Calo-Mata P, Villa TG (June 2017). "Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria". Biochemical Pharmacology. 133 (6): 117–138. doi:10.1016/j.bcp.2016.09.018. PMID 27663838.
- Reddy KV, Yedery RD, Aranha C (December 2004). "Antimicrobial peptides: premises and promises". International Journal of Antimicrobial Agents. 24 (6): 536–547. doi:10.1016/j.ijantimicag.2004.09.005. PMID 15555874.
- Yeaman MR, Yount NY (March 2003). "Mechanisms of antimicrobial peptide action and resistance". Pharmacological Reviews. 55 (1): 27–55. doi:10.1124/pr.55.1.2. PMID 12615953. S2CID 6731487.
- Papagianni M (September 2003). "Ribosomally synthesized peptides with antimicrobial properties: biosynthesis, structure, function, and applications". Biotechnology Advances. 21 (6): 465–499. doi:10.1016/S0734-9750(03)00077-6. PMID 14499150.
- Sitaram N, Nagaraj R (2002). "Host-defense antimicrobial peptides: importance of structure for activity". Current Pharmaceutical Design. 8 (9): 727–742. doi:10.2174/1381612023395358. PMID 11945168.
- Dürr UH, Sudheendra US, Ramamoorthy A (September 2006). "LL-37, the only human member of the cathelicidin family of antimicrobial peptides". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1758 (9): 1408–1425. doi:10.1016/j.bbamem.2006.03.030. PMID 16716248.
- Dhople V, Krukemeyer A, Ramamoorthy A (September 2006). "The human beta-defensin-3, an antibacterial peptide with multiple biological functions". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1758 (9): 1499–1512. doi:10.1016/j.bbamem.2006.07.007. PMID 16978580.
- Hancock RE, Rozek A (January 2002). "Role of membranes in the activities of antimicrobial cationic peptides". FEMS Microbiology Letters. 206 (2): 143–149. doi:10.1111/j.1574-6968.2002.tb11000.x. PMID 11814654.
- Varkey J, Singh S, Nagaraj R (November 2006). "Antibacterial activity of linear peptides spanning the carboxy-terminal beta-sheet domain of arthropod defensins". Peptides. 27 (11): 2614–2623. doi:10.1016/j.peptides.2006.06.010. PMID 16914230. S2CID 21104756.
- Nguyen LT, Haney EF, Vogel HJ (September 2011). "The expanding scope of antimicrobial peptide structures and their modes of action". Trends in Biotechnology. 29 (9): 464–472. doi:10.1016/j.tibtech.2011.05.001. PMID 21680034.
- O'Driscoll NH, Labovitiadi O, Cushnie TP, Matthews KH, Mercer DK, Lamb AJ (March 2013). "Production and evaluation of an antimicrobial peptide-containing wafer formulation for topical application". Current Microbiology. 66 (3): 271–278. doi:10.1007/s00284-012-0268-3. PMID 23183933. S2CID 18671683.
- Hein KZ, Takahashi H, Tsumori T, Yasui Y, Nanjoh Y, Toga T, et al. (October 2015). "Disulphide-reduced psoriasin is a human apoptosis-inducing broad-spectrum fungicide". Proceedings of the National Academy of Sciences of the United States of America. 112 (42): 13039–13044. Bibcode:2015PNAS..11213039H. doi:10.1073/pnas.1511197112. PMC 4620902. PMID 26438863.
- Brogden KA (March 2005). "Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria?". Nature Reviews. Microbiology. 3 (3): 238–250. doi:10.1038/nrmicro1098. PMID 15703760. S2CID 23625167.
- Hirst DJ, Lee TH, Swann MJ, Unabia S, Park Y, Hahm KS, Aguilar MI (April 2011). "Effect of acyl chain structure and bilayer phase state on binding and penetration of a supported lipid bilayer by HPA3". European Biophysics Journal. 40 (4): 503–514. doi:10.1007/s00249-010-0664-1. PMID 21222117. S2CID 22514308.
- Lee TH, Heng C, Swann MJ, Gehman JD, Separovic F, Aguilar MI (October 2010). "Real-time quantitative analysis of lipid disordering by aurein 1.2 during membrane adsorption, destabilisation and lysis". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1798 (10): 1977–1986. doi:10.1016/j.bbamem.2010.06.023. PMID 20599687.
- Turnidge JD, Bell JM (1996). "Susceptibility testing of Antimicrobials in liquid media". In Lorian V (ed.). Antibiotics in Laboratory Medicine (4th ed.). Baltimore, Md.: Williams and Wilkins. pp. 52–111. ISBN 978-0-683-05169-8.
National Committee of Laboratory Safety and Standards (NCLSS)
- Wang CK, Shih LY, Chang KY (November 2017). "Large-Scale Analysis of Antimicrobial Activities in Relation to Amphipathicity and Charge Reveals Novel Characterization of Antimicrobial Peptides". Molecules. 22 (11): 2037. doi:10.3390/molecules22112037. PMC 6150348. PMID 29165350.
- Hunter HN, Fulton DB, Ganz T, Vogel HJ (October 2002). "The solution structure of human hepcidin, a peptide hormone with antimicrobial activity that is involved in iron uptake and hereditary hemochromatosis". The Journal of Biological Chemistry. 277 (40): 37597–37603. doi:10.1074/jbc.M205305200. PMID 12138110.
- Kudrimoti M, Curtis A, Azawi S, Worden F, Katz S, Adkins D, et al. (December 2016). "Dusquetide: A novel innate defense regulator demonstrating a significant and consistent reduction in the duration of oral mucositis in preclinical data and a randomized, placebo-controlled phase 2a clinical study". Journal of Biotechnology. 239: 115–125. doi:10.1016/j.jbiotec.2016.10.010. PMID 27746305.
- Guilhelmelli F, Vilela N, Albuquerque P, Derengowski L, Silva-Pereira I, Kyaw CM (December 2013). "Antibiotic development challenges: the various mechanisms of action of antimicrobial peptides and of bacterial resistance". Frontiers in Microbiology. 4 (4): 353. doi:10.3389/fmicb.2013.00353. PMC 3856679. PMID 24367355.
- Hallock KJ, Lee DK, Ramamoorthy A (May 2003). "MSI-78, an analogue of the magainin antimicrobial peptides, disrupts lipid bilayer structure via positive curvature strain". Biophysical Journal. 84 (5): 3052–3060. Bibcode:2003BpJ....84.3052H. doi:10.1016/S0006-3495(03)70031-9. PMC 1302867. PMID 12719236.
- Henzler Wildman KA, Lee DK, Ramamoorthy A (June 2003). "Mechanism of lipid bilayer disruption by the human antimicrobial peptide, LL-37". Biochemistry. 42 (21): 6545–6558. doi:10.1021/bi0273563. PMID 12767238.
- Järvå M, Lay FT, Phan TK, Humble C, Poon IK, Bleackley MR, et al. (May 2018). "X-ray structure of a carpet-like antimicrobial defensin-phospholipid membrane disruption complex". Nature Communications. 9 (1): 1962. Bibcode:2018NatCo...9.1962J. doi:10.1038/s41467-018-04434-y. PMC 5958116. PMID 29773800.
- Poon IK, Baxter AA, Lay FT, Mills GD, Adda CG, Payne JA, et al. (April 2014). "Phosphoinositide-mediated oligomerization of a defensin induces cell lysis". eLife. 3: e01808. doi:10.7554/elife.01808. PMC 3968744. PMID 24692446.
- Järvå M, Phan TK, Lay FT, Caria S, Kvansakul M, Hulett MD (July 2018). "Human β-defensin 2 kills Candida albicans through phosphatidylinositol 4,5-bisphosphate-mediated membrane permeabilization". Science Advances. 4 (7): eaat0979. Bibcode:2018SciA....4..979J. doi:10.1126/sciadv.aat0979. PMC 6059731. PMID 30050988.
- Balatti GE, Ambroggio EE, Fidelio GD, Martini MF, Pickholz M (October 2017). "Differential Interaction of Antimicrobial Peptides with Lipid Structures Studied by Coarse-Grained Molecular Dynamics Simulations". Molecules. 22 (10): 1775. doi:10.3390/molecules22101775. PMC 6151434. PMID 29053635.
- Gomes B, Augusto MT, Felício MR, Hollmann A, Franco OL, Gonçalves S, Santos NC (9 January 2018). "Designing improved active peptides for therapeutic approaches against infectious diseases". Biotechnology Advances. 36 (2): 415–429. doi:10.1016/j.biotechadv.2018.01.004. PMID 29330093.
- Hanson MA, Lemaitre B (February 2020). "New insights on Drosophila antimicrobial peptide function in host defense and beyond". Current Opinion in Immunology. 62: 22–30. doi:10.1016/j.coi.2019.11.008. PMID 31835066.
- Mookherjee N, Anderson MA, Haagsman HP, Davidson DJ (May 2020). "Antimicrobial host defence peptides: functions and clinical potential" (PDF). Nature Reviews. Drug Discovery. 19 (5): 311–332. doi:10.1038/s41573-019-0058-8. hdl:20.500.11820/1ec38809-e8f2-4684-8bbb-e908ecb5c66e. PMID 32107480. S2CID 211526624.
- Hoskin DW, Ramamoorthy A (February 2008). "Studies on anticancer activities of antimicrobial peptides". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1778 (2): 357–375. doi:10.1016/j.bbamem.2007.11.008. PMC 2238813. PMID 18078805.
- Parvy JP, Yu Y, Dostalova A, Kondo S, Kurjan A, Bulet P, et al. (July 2019). "The antimicrobial peptide defensin cooperates with tumour necrosis factor to drive tumour cell death in Drosophila". eLife. 8. doi:10.7554/eLife.45061. PMC 6667213. PMID 31358113.
- Kalsy M, Tonk M, Hardt M, Dobrindt U, Zdybicka-Barabas A, Cytrynska M, et al. (February 2020). "The insect antimicrobial peptide cecropin A disrupts uropathogenic Escherichia coli biofilms". NPJ Biofilms and Microbiomes. 6 (1): 6. doi:10.1038/s41522-020-0116-3. PMC 7016129. PMID 32051417.
- Hassan M, Kjos M, Nes IF, Diep DB, Lotfipour F (October 2012). "Natural antimicrobial peptides from bacteria: characteristics and potential applications to fight against antibiotic resistance". Journal of Applied Microbiology. 113 (4): 723–736. doi:10.1111/j.1365-2672.2012.05338.x. PMID 22583565.
- Kumaresan V, Bhatt P, Ganesh MR, Harikrishnan R, Arasu M, Al-Dhabi NA, et al. (December 2015). "A novel antimicrobial peptide derived from fish goose type lysozyme disrupts the membrane of Salmonella enterica". Molecular Immunology. 68 (2 Pt B): 421–433. doi:10.1016/j.molimm.2015.10.001. PMID 26477736.
- Arasu A, Kumaresan V, Ganesh MR, Pasupuleti M, Arasu MV, Al-Dhabi NA, Arockiaraj J (June 2017). "Bactericidal activity of fish galectin 4 derived membrane-binding peptide tagged with oligotryptophan". Developmental and Comparative Immunology. 71: 37–48. doi:10.1016/j.dci.2017.01.019. PMID 28126555. S2CID 22374102.
- Arockiaraj J, Chaurasia MK, Kumaresan V, Palanisamy R, Harikrishnan R, Pasupuleti M, Kasi M (April 2015). "Macrobrachium rosenbergii mannose binding lectin: synthesis of MrMBL-N20 and MrMBL-C16 peptides and their antimicrobial characterization, bioinformatics and relative gene expression analysis". Fish & Shellfish Immunology. 43 (2): 364–374. doi:10.1016/j.fsi.2014.12.036. PMID 25575476.
- Somasekhar M (24 April 2019). "In Spiny anteater's milk could be an alternative to antibiotics". BusinessLine. Chennai, India: Kasturi & Sons Ltd. Retrieved 17 January 2021.
{{cite news}}: CS1 maint: url-status (link) - "Egg-laying mammal's milk may help for novel antibiotics". Deccan Herald. India: The Printers Mysore. Press Trust of India. 25 April 2019. Retrieved 17 January 2021.
{{cite news}}: CS1 maint: url-status (link) - Matsuzaki K (August 2009). "Control of cell selectivity of antimicrobial peptides". Biochimica et Biophysica Acta (BBA) - Biomembranes (Review article). 1788 (8): 1687–1692. doi:10.1016/j.bbamem.2008.09.013. PMID 18952049.
- Sawaki K, Mizukawa N, Yamaai T, Yoshimoto T, Nakano M, Sugahara T (2002). "High concentration of beta-defensin-2 in oral squamous cell carcinoma". Anticancer Research. 22 (4): 2103–2107. PMID 12174890.
- Kida-Takaoka S, Yamaai T, Mizukawa N, Murakami J, Iida S (November 2014). "Surrounding cells affect the gene expression pattern of human beta-defensins in squamous cell carcinoma in vitro". Anticancer Research. 34 (11): 6443–6449. PMID 25368244. Retrieved 7 February 2019.
- Hancock RE, Sahl HG (December 2006). "Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies". Nature Biotechnology (Review article). 24 (12): 1551–1557. doi:10.1038/nbt1267. PMID 17160061. S2CID 22384207.
- Zasloff M (January 2002). "Antimicrobial peptides of multicellular organisms". Nature (Review article). 415 (6870): 389–395. Bibcode:2002Natur.415..389Z. doi:10.1038/415389a. PMID 11807545. S2CID 205028607.
- Matsuzaki K, Sugishita K, Fujii N, Miyajima K (March 1995). "Molecular basis for membrane selectivity of an antimicrobial peptide, magainin 2". Biochemistry. 34 (10): 3423–3429. doi:10.1021/bi00010a034. PMID 7533538.
- Chou HT, Kuo TY, Chiang JC, Pei MJ, Yang WT, Yu HC, et al. (August 2008). "Design and synthesis of cationic antimicrobial peptides with improved activity and selectivity against Vibrio spp". International Journal of Antimicrobial Agents. 32 (2): 130–138. doi:10.1016/j.ijantimicag.2008.04.003. PMID 18586467.
- Tennessen JA (November 2005). "Molecular evolution of animal antimicrobial peptides: widespread moderate positive selection". Journal of Evolutionary Biology (Review article). 18 (6): 1387–1394. doi:10.1111/j.1420-9101.2005.00925.x. PMID 16313451.
- Yu L, Guo L, Ding JL, Ho B, Feng SS, Popplewell J, et al. (February 2009). "Interaction of an artificial antimicrobial peptide with lipid membranes". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1788 (2): 333–344. doi:10.1016/j.bbamem.2008.10.005. PMID 19013127.
- Lee TH, Hall K, Mechler A, Martin L, Popplewell J, Ronan G, Aguilar MI (2009). "Molecular imaging and orientational changes of antimicrobial peptides in membranes". Peptides for Youth. Advances in Experimental Medicine and Biology. Vol. 611. pp. 313–315. Bibcode:2009peyo.book..313L. doi:10.1007/978-0-387-73657-0_140. ISBN 978-0-387-73656-3. PMID 19400207.
- Zhu WL, Lan H, Park Y, Yang ST, Kim JI, Park IS, et al. (October 2006). "Effects of Pro --> peptoid residue substitution on cell selectivity and mechanism of antibacterial action of tritrpticin-amide antimicrobial peptide". Biochemistry. 45 (43): 13007–13017. doi:10.1021/bi060487+. PMID 17059217.
- Kulagina NV, Lassman ME, Ligler FS, Taitt CR (October 2005). "Antimicrobial peptides for detection of bacteria in biosensor assays". Analytical Chemistry. 77 (19): 6504–6508. doi:10.1021/ac050639r. PMID 16194120.
- Peschel A, Otto M, Jack RW, Kalbacher H, Jung G, Götz F (March 1999). "Inactivation of the dlt operon in Staphylococcus aureus confers sensitivity to defensins, protegrins, and other antimicrobial peptides". The Journal of Biological Chemistry. 274 (13): 8405–8410. doi:10.1074/jbc.274.13.8405. PMID 10085071.
- Campos MA, Vargas MA, Regueiro V, Llompart CM, Albertí S, Bengoechea JA (December 2004). Weiser JN (ed.). "Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides". Infection and Immunity. 72 (12): 7107–7114. doi:10.1128/IAI.72.12.7107-7114.2004. PMC 529140. PMID 15557634.
- China B, N'Guyen BT, de Bruyere M, Cornelis GR (April 1994). "Role of YadA in resistance of Yersinia enterocolitica to phagocytosis by human polymorphonuclear leukocytes". Infection and Immunity. 62 (4): 1275–1281. doi:10.1128/IAI.62.4.1275-1281.1994. PMC 186269. PMID 8132334.
- Shelton CL, Raffel FK, Beatty WL, Johnson SM, Mason KM (November 2011). "Sap transporter mediated import and subsequent degradation of antimicrobial peptides in Haemophilus". PLOS Pathogens. 7 (11): e1002360. doi:10.1371/journal.ppat.1002360. PMC 3207918. PMID 22072973.
- Nikaido H (October 1996). "Multidrug efflux pumps of gram-negative bacteria". Journal of Bacteriology. 178 (20): 5853–5859. doi:10.1128/jb.178.20.5853-5859.1996. PMC 178438. PMID 8830678.
- Whitelock JM, Murdoch AD, Iozzo RV, Underwood PA (April 1996). "The degradation of human endothelial cell-derived perlecan and release of bound basic fibroblast growth factor by stromelysin, collagenase, plasmin, and heparanases". The Journal of Biological Chemistry. 271 (17): 10079–10086. doi:10.1074/jbc.271.17.10079. PMID 8626565.
- Kulkarni HM, Swamy C, Jagannadham MV (March 2014). "Molecular characterization and functional analysis of outer membrane vesicles from the antarctic bacterium Pseudomonas syringae suggest a possible response to environmental conditions". Journal of Proteome Research. 13 (3): 1345–1358. doi:10.1021/pr4009223. PMID 24437924.
- Chua SL, Tan SY, Rybtke MT, Chen Y, Rice SA, Kjelleberg S, et al. (May 2013). "Bis-(3'-5')-cyclic dimeric GMP regulates antimicrobial peptide resistance in Pseudomonas aeruginosa". Antimicrobial Agents and Chemotherapy. 57 (5): 2066–2075. doi:10.1128/AAC.02499-12. PMC 3632963. PMID 23403434.
- Habets MG, Brockhurst MA (June 2012). "Therapeutic antimicrobial peptides may compromise natural immunity". Biology Letters. 8 (3): 416–418. doi:10.1098/rsbl.2011.1203. PMC 3367763. PMID 22279153.
- Bahar AA, Ren D (November 2013). "Antimicrobial peptides". Pharmaceuticals. 6 (12): 1543–1575. doi:10.3390/ph6121543. PMC 3873676. PMID 24287494.
- Gumienna-Kontecka E, Carver PL (2019). "Chapter 7. Building a Trojan Horse: Siderophore-Drug Conjugates for the Treatment of Infectious Diseases". In Sigel A, Freisinger E, Sigel RK, Carver PL (eds.). Essential Metals in Medicine:Therapeutic Use and Toxicity of Metal Ions in the Clinic. Metal Ions in Life Sciences. Vol. 19. Berlin: de Gruyter GmbH. pp. 181–202. doi:10.1515/9783110527872-013. ISBN 978-3-11-052691-2. PMID 30855108.
- "Insect antimicrobial peptides OPM". Orientations of Proteins in Membranes database (OPM).
- Szymanowski F, Balatti GE, Ambroggio E, Hugo AA, Martini MF, Fidelio GD, et al. (June 2019). "Differential activity of lytic α-helical peptides on lactobacilli and lactobacilli-derived liposomes". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1861 (6): 1069–1077. doi:10.1016/j.bbamem.2019.03.004. PMID 30878358.
- "Amphibian antimicrobial peptides". Orientations of Proteins in Membranes database (OPM).
- Yang M, Zhang C, Zhang X, Zhang MZ, Rottinghaus GE, Zhang S (September 2016). "Structure-function analysis of Avian β-defensin-6 and β-defensin-12: role of charge and disulfide bridges". BMC Microbiology. 16: 210. doi:10.1186/s12866-016-0828-y. PMC 5016922. PMID 27613063.
- Niedermaier H (9 February 2012). "Synthetic Mimics of Antimicrobial Peptides: A New Wave of Antibiotics". Department of Chemistry. University of Georgia. Archived from the original on 28 October 2016. Retrieved 27 October 2016.
- Kohn EM, Shirley DJ, Arotsky L, Picciano AM, Ridgway Z, Urban MW, et al. (February 2018). "Role of Cationic Side Chains in the Antimicrobial Activity of C18G". Molecules. 23 (2): 329. doi:10.3390/molecules23020329. PMC 6017431. PMID 29401708.
- Shin JM, Gwak JW, Kamarajan P, Fenno JC, Rickard AH, Kapila YL (June 2016). "Biomedical applications of nisin". Journal of Applied Microbiology. 120 (6): 1449–1465. doi:10.1111/jam.13033. PMC 4866897. PMID 26678028.
- Lee HT, Lee CC, Yang JR, Lai JZ, Chang KY (February 2015). "A large-scale structural classification of antimicrobial peptides". BioMed Research International. 2015: 475062. doi:10.1155/2015/475062. PMC 4426897. PMID 26000295.
- Waghu FH, Gopi L, Barai RS, Ramteke P, Nizami B, Idicula-Thomas S (January 2014). "CAMP: Collection of sequences and structures of antimicrobial peptides". Nucleic Acids Research. 42 (Database issue): D1154–D1158. doi:10.1093/nar/gkt1157. PMC 3964954. PMID 24265220.
- Shi GB, Kang XY, Dong FY, Liu YC, Zhu N (January 2022). "DRAMP 3.0: an enhanced comprehensive data repository of antimicrobial peptides". Nucleic Acids Research. 50 (Database issue): D488–D496. doi:10.1093/nar/gkab651. PMC 8728287. PMID 34390348.
- Jhong JH, Chi YH, Li WC, Lin TH, Huang KY, Lee TY (January 2019). "dbAMP: an integrated resource for exploring antimicrobial peptides with functional activities and physicochemical properties on transcriptome and proteome data". Nucleic Acids Research. 47 (D1): D285–D297. doi:10.1093/nar/gky1030. PMC 6323920. PMID 30380085.
- Mooney C, Haslam NJ, Pollastri G, Shields DC (8 October 2012). "Towards the improved discovery and design of functional peptides: common features of diverse classes permit generalized prediction of bioactivity". PLOS ONE. 7 (10): e45012. Bibcode:2012PLoSO...745012M. doi:10.1371/journal.pone.0045012. PMC 3466233. PMID 23056189.
- Mooney C, Haslam NJ, Holton TA, Pollastri G, Shields DC (May 2013). "PeptideLocator: prediction of bioactive peptides in protein sequences". Bioinformatics. 29 (9): 1120–1126. doi:10.1093/bioinformatics/btt103. PMID 23505299.
- Müller AT, Gabernet G, Hiss JA, Schneider G (September 2017). "modlAMP: Python for antimicrobial peptides". Bioinformatics. 33 (17): 2753–2755. doi:10.1093/bioinformatics/btx285. PMID 28472272.
- Agrawal P, Raghava GP (26 October 2018). "Prediction of Antimicrobial Potential of a Chemically Modified Peptide From Its Tertiary Structure". Frontiers in Microbiology. 9: 2551. doi:10.3389/fmicb.2018.02551. PMC 6212470. PMID 30416494.
- Agrawal P, Bhalla S, Chaudhary K, Kumar R, Sharma M, Raghava GP (26 February 2018). "In Silico Approach for Prediction of Antifungal Peptides". Frontiers in Microbiology. 9: 323. doi:10.3389/fmicb.2018.00323. PMC 5834480. PMID 29535692.
- Usmani SS, Bhalla S, Raghava GP (26 August 2018). "Prediction of Antitubercular Peptides From Sequence Information Using Ensemble Classifier and Hybrid Features". Frontiers in Pharmacology. 9: 954. doi:10.3389/fphar.2018.00954. PMC 6121089. PMID 30210341.
External links
- ADAM (A Database of Anti-Microbial peptides) at ntou.edu.tw
- AntiFP Prediction of antifungal peptides
- AntiMPmod Prediction of antimicrobial potential of modified peptides
- Antimicrobial+Cationic+Peptides at the US National Library of Medicine Medical Subject Headings (MeSH)
- AntiTbPred Prediction of anti-tuberculosis peptides
- Antimicrobial Peptide Database at University of Nebraska Medical Center
- Antimicrobial Peptide Scanner Deep Learning based AMP prediction server
- AntiTbPdb Anti Tubercular Peptide Database
- BioPD at Peking University Health Science Center
- CAMP:Collection of Anti-Microbial Peptides at National Institute for Research in Reproductive Health (NIRRH)
- DBAASP - Database of Antimicrobial Activity and Structure of Peptides]
- LAMP at Fudan University
- PeptideLocator Prediction of functional peptides, including antimicrobial peptides, in a protein sequence
- PeptideRanker Bioactive peptide, including antimicrobial peptide, prediction
- modlAMP Python package for computational work with antimicrobial peptides, including sequence handling, -design, -prediction, descriptor calculation and plotting