Central pattern generator
Central pattern generators (CPGs) are self-organizing biological neural circuits[1][2][3] that produce rhythmic outputs in the absence of rhythmic input.[4][5][6] They are the source of the tightly-coupled patterns of neural activity that drive rhythmic and stereotyped motor behaviors like walking, swimming, breathing, or chewing. The ability to function without input from higher brain areas still requires modulatory inputs, and their outputs are not fixed. Flexibility in response to sensory input is a fundamental quality of CPG-driven behavior.[4][5] To be classified as a rhythmic generator, a CPG requires:
- "two or more processes that interact such that each process sequentially increases and decreases, and
- that, as a result of this interaction, the system repeatedly returns to its starting condition."[4]
CPGs have been found in invertebrates,[7] and practically all vertebrate species investigated,[8][9] including humans.[10][11][12]
General anatomy and physiology
Intrinsic properties of CPG neurons
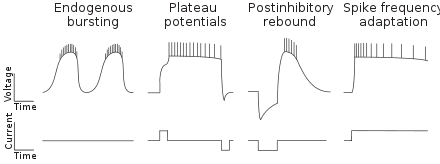
CPG neurons can have different intrinsic membrane properties (see schematic).[13] Some neurons fire bursts of action potentials, either endogenously or in the presence of neuromodulatory substances. Other neurons are bistable and generate plateau potentials that can be triggered by a depolarizing current pulse, and terminated by a hyperpolarizing current pulse. Many CPG neurons fire after being released from inhibition (postinhibitory rebound). Another common feature of CPG neurons is a decrease in the frequency of firing during a constant depolarization (spike frequency adaptation).
Mechanisms of rhythm generation
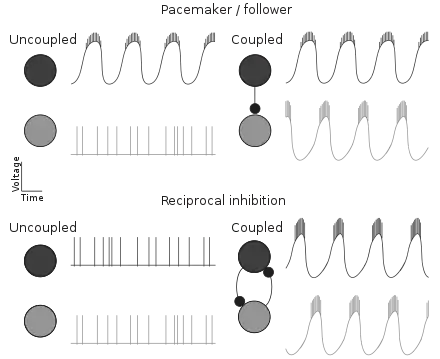
Rhythm generation in CPG networks depends on the intrinsic properties of CPG neurons and their synaptic connections. There are two general mechanisms for rhythm generation: pacemaker/follower and reciprocal inhibition (see schematic).[13]
In a network driven by a pacemaker, one or more neurons act as a core oscillator (pacemaker) that drives other, non-bursting neurons (follower) into a rhythmic pattern. Examples of pacemaker driven networks include the pyloric rhythm of the crustacean stomatogastric ganglion[14] and the vertebrate respiratory rhythms.[15]
In a network driven by reciprocal inhibition, two (groups of) neurons reciprocally inhibit each other. Such networks are known as half-center oscillators. The neurons are not rhythmically active when isolated, but they can produce alternating patterns of activity when coupled by inhibitory connections. (The neurons can also produce activity patterns of other relative phasing, including synchrony, depending on the synaptic properties). The transitions between activated and inhibited states can occur via a number of mechanisms. For example, spike-frequency adaptation in the bursting neuron(s) may slowly release the other neuron(s) from inhibition.[16] Reciprocal inhibition is a core feature of many CPGs, including those involved in locomotion.[17][18][19][20]
Short-term synaptic dynamics in CPG networks
CPG networks have extensive recurrent synaptic connections including reciprocal excitation and reciprocal inhibition. Synapses in CPG networks are subject to short-term activity dependent modifications.[21][22] Short-term synaptic depression and facilitation of synapses can play a role in transitions between active and inactive phases of bursting and termination of bursts.[23][24]
Location and identity of CPG neurons
CPG circuits thought to be involved in the control of locomotion consist of motor neurons and spinal interneurons and are located in the lower thoracic and lumbar regions of the vertebrate spinal cord ,[25] and in each neuromere of the invertebrate ventral nerve cord.[26] CPG neurons involved in swallowing are located in the brain stem, specifically the hypoglossal nucleus within the medulla.[27][28]
While the general location of CPG neurons can often be inferred, the specific location and identity of the participating neurons are only beginning to be understood. CPG networks are often distributed and can be flexibly reorganized, making it challenging to identify spinal interneurons. Over the past few decades, molecular and genetic programs that control neuronal patterning have been used to specifically target spinal interneurons in mice[29] and zebrafish.[30] Developing neural tube of embryonic mouse shows expression of distinct transcription factors in domains along the dorso-ventral axis of the spinal cord.[31] These domains give rise to distinct population of neurons that have been classified as dorsal (dI1-dI6) and ventral (V0-V3) cardinal classes of spinal interneurons.[32][33][29] The ventral neurons are considered to be members of the spinal CPG network.[29] Each of these interneuron class can be further divided into diverse subpopulations of neurons with distinct neurotransmitter phenotype, axonal projection and function during locomotion.[32] For example, V2 interneurons are ipsilaterally projecting that can be further classified as excitatory V2a and inhibitory V2b. V2s are important for flexor-extensor alternation and provide excitation to motor neurons.[32]
Neuromodulation
Organisms must adapt their behavior to meet the needs of their internal and external environments. Central pattern generators, as part of the neural circuitry of an organism, can be modulated to adapt to the organism's needs and surroundings. Three roles of neuromodulation have been found for CPG circuits:[4]
- Modulation is intrinsic to CPG network or required for its activation
- Modulation changes the functional configuration of CPGs to produce different outputs
- Modulation alters CPG neuron complement by switching neurons between networks and fusing formerly separate networks into larger entities
Modulation is intrinsic to CPG network or required for its activation
Neuromodulatory synapses may be part of the CPG network itself.[34] For example, the CPG underlying the escape swim response in Tritonia diomedea contains intrinsic neuromodulatory neurons. These neuromodulatory neurons can enhance neurotransmitter release from another neuron in the circuit and its neuromodulatory actions are thought to be important for producing swim motor program.[35] Neuromodulatory inputs can also activate CPG network and can be necessary for generation of rhythmic output. Loss of neuromodulatory inputs can abolish rhythmic activity from pyloric network.[34] In vertebrates, application of neuromodulators have been shown to evoke locomotor activity.[36]
Modulation changes the functional configuration of CPGs to produce different outputs
Neuromodulators can alter the synaptic strength as well as intrinsic properties of neurons.[37] These actions can alter the frequency and the phase relationships between neurons and thereby change the output pattern of the circuit. For example, exogenous application of various neuromodulators can elicit triphasic motor pattern in the STG where each modulator results in generation of a different motor pattern.[38] Neuromodulatory projections expressing a common modulator can also elicit different patterns from the same network. Stimulation of different proctolin-containing projection neurons in the STG results in a distinct motor pattern from the same network due to differences in cotransmitter complement of these projection neurons.[39]
The effects of neuromodulators are distributed throughout the CPG network. Specially, dopamine was shown to affect cellular and synaptic properties of nearly all components of the crustacean pyloric network. Moreover, dopamine can have opposing effects on different components of the network. As such, the final output of the network reflects a combination of modulatory actions on individual components.[40]
Modulation alters CPG neuron complement by switching neurons between networks and fusing formerly separate networks into larger entities.
Neuronal composition of CPGs can vary with the state of the system.[41] Neuromodulators can activate or inhibit neurons of the CPGs and can even combine different networks into one.[38] For example, in the lobster stomatogastric nervous system the neuropeptide, red pigment concentrating hormone, can strengthen synapses between two different networks to create a single, combined rhythm.[42] Neuromodulators can also result in switching of neurons from one network to another.[43]
Sensory feedback
Although the theory of central pattern generation calls for basic rhythmicity and patterning to be centrally generated, CPGs can respond to sensory feedback to alter the patterning in behaviorally appropriate ways. Alteration of the pattern is difficult because feedback received during only one phase may require changed movement in the other parts of the patterned cycle to preserve certain coordination relationships. For example, walking with a pebble in the right shoe alters the entire gait, even though the stimulus is only present while standing on the right foot. Even during the time when the left foot is down and the sensory feedback is inactive, action is taken to prolong the right leg swing and extend the time on the left foot, leading to limping. This effect could be due to widespread and long-lasting effects of the sensory feedback on the CPG or due to short-term effects on a few neurons that in turn modulate nearby neurons and spread the feedback through the entire CPG in that way. Some degree of modulation is required to allow one CPG to assume multiple states in response to feedback.[4]
Additionally, the effect of the sensory input varies depending on the phase of the pattern in which it occurs. For example, during walking, resistance to the top of the swinging foot (e.g., by a horizontal stick) causes the foot to be lifted higher to move over the stick. However, the same input to the standing foot cannot cause the foot to lift or the person would collapse. Thus, depending on the phase, the same sensory input can cause the foot to be lifted higher or held more firmly to the ground. "This change in motor response as a function of motor pattern phase is called reflex reversal, and has been observed in invertebrates (DiCaprio and Clarac, 1981) and vertebrates (Forssberg et al., 1977). How this process occurs is poorly understood, but again two possibilities exist. One is that sensory input is appropriately routed to different CPG neurons as a function of motor pattern phase. The other is that the input reaches the same neurons at all phases, but that, as a consequence of the way in which the network transforms the input, network response varies appropriately as a function of motor pattern phase."[4]
A study by Gottschall and Nichols examined the hindlimb of a decerebrate cat during walking (a CPG controlled function) in response to changes in head pitch. This study describes the differences in gait and body position of cats walking uphill, downhill and on level surfaces. Proprioceptive (Golgi tendon organs and muscle spindles) and exteroreceptive (optic, vestibular and cutaneous) receptors work alone or in combination to adjust the CPG to sensory feedback. The study explored the effects of neck proprioceptors (giving information about the relative location of the head and body) and vestibular receptors (giving information about the orientation of the head relative to gravity). Decerebrate cats were made to walk on a level surface with their heads level, tilted up or tilted down. Comparing the decerebrate cats to normal cats showed similar EMG patterns during level walking and EMG patterns that reflected downhill walking with the head titled up and uphill walking with the head tilted down. This study proved that neck proprioceptors and vestibular receptors contribute sensory feedback that alters the gait of the animal. This information may be useful for treatment of gait disorders.[44]
Functions
Central pattern generators can serve many functions. CPGs can play roles in movement, breathing, rhythm generation and other oscillatory functions. Below are several key functions of CPGs.
Locomotion
As early as 1911, it was recognized, by the experiments of Thomas Graham Brown, that the basic pattern of stepping can be produced by the spinal cord without the need of descending commands from the cortex.[45][46]
The first modern evidence of the central pattern generator was produced by isolating the locust nervous system and showing that it could produce a rhythmic output in isolation resembling that of the locust in flight. This was discovered by Wilson in 1961.[4] Since that time, evidence has arisen for the presence of central pattern generators in vertebrate animals, starting with work on the cat in the 1960s by Elzbieta Jankowska in Gothenburg, who provided the first evidence for a spinal cord CPG. This section addresses the role of the central pattern generator in locomotion for the lamprey and humans.
The lamprey has been used as a model for vertebrate CPGs because, while its nervous system has a vertebrate organization, it shares many positive characteristics with invertebrates. When removed from the lamprey, the intact spinal cord can survive for days in vitro. It also has very few neurons and can be easily stimulated to produce a fictive swimming motion indicative of a central pattern generator. As early as 1983, Ayers, Carpenter, Currie and Kinch proposed that there was a CPG responsible for most undulating movements in the lamprey including swimming forward and backward, burrowing in the mud and crawling on a solid surface, that although not surprisingly did not match the activity in the intact animal, nevertheless provided the basic locomotor output.[47] The different movements have been found to be altered by neuromodulators, including serotonin in a study by Harris-Warrick and Cohen in 1985[48] and tachykinin in a study by Parker et al.[49] in 1998. The lamprey model of CPG for locomotion has been important to the study of CPGs. Although Sten Grillner claims that the locomotor network is characterised, a claim that has seemingly been uncritically accepted by the spinal cord locomotor network field, there are in fact many missing details and Grillner cannot provide the evidence he uses to support his claims (Parker 2006).[50][51] However, this neural circuit model[52] of the lamprey CPG, including three classes (one excitatory and two inhibitory) of neurons but omitting sub-cellular details, provides a system level understanding of the CPG-generated locomotion whose speed and direction (swimming forward, backward, or turning) are set by non-rhythmic external inputs (from the brainstem) to the circuit.[52] A general scheme of the lamprey CPG is now being used in the creation of artificial CPGs. For example, Ijspeert and Kodjabachian used Ekeberg's model for the lamprey to create artificial CPGs and simulate swimming movements in a lamprey-like substrate using controllers based on a SGOCE encoding.[53] Essentially, these are the first steps toward the use of CPGs to code for locomotion in robots. The vertebrate model of CPG has been also developed with both Hodgkin-Huxley formalism,[54] its variants [55] and control system approaches.[56][57] For example, Yakovenko and colleagues have developed a simple mathematical model that describes basic principles proposed by T.G. Brown with integrate-to-threshold units organized with mutually inhibitory connections. This model is sufficient to describe complex properties of behavior, such as different regimes of the extensor- and flexor-dominant locomotion observed during electrical stimulation of the mesencephalic locomotor region (MLR), MLR-induced fictive locomotion.[57]
The connections between the CPGs that control each limb manage interlimb coordination and hence the gaits in quadrupedal and possibly also bipedal animals.[58][59][60][61][62] Left right coordination is mediated by commissural and fore-hind as well as diagonal coordination is mediated by long-projecting propiospinal interneurons.[63][64] The balance of the left-right alternation (mediated genetically identified V0d and V0v neuron classes) to left-synchronization promoting commissural interneurons (potentially mediated V3 neurons) determines whether walk and trot (alternating gaits) or gallop and bound (synchronous gaits) are expressed.[58] This balance changes with increasing speed, potentially because of modulation by supraspinal drive from the MLR and mediated by the reticular formation, and causes speed dependent gait transitions characteristic for quadrupedal animals.[58][61][65] The walk to trot transition potentially occurs because of the stronger decrease of extension than flexion phase durations with increasing locomotor speed and could be mediated by descending diagonal inhibition through V0d long propriospinal neurons,[61] which leads to progressively increasing overlap between the diagonal limbs up until diagonal synchronization (trot).[58] Commissural and long propriospinal neurons are a likely target of supraspinal and somatosensory afferent inputs to adjust interlimb coordination and gait to different environmental and behavioral conditions.[61]
Central pattern generators also contribute to locomotion in humans. In 1994, Calancie, et al. described the "first well-defined example of a central rhythm generator for stepping in the adult human." The subject was a 37-year-old male who suffered an injury to the cervical spinal cord 17 years prior. After initial total paralysis below the neck, the subject eventually regained some movement of the arms and fingers and limited movement in the lower limbs. He had not recovered sufficiently to support his own weight. After 17 years, the subject found that when lying supine and extending his hips, his lower extremities underwent step-like movements for as long as he remained lying down. "The movements (i) involved alternating flexion and extension of his hips, knees, and ankles; (ii) were smooth and rhythmic; (iii) were forceful enough that the subject soon became uncomfortable due to excessive muscle 'tightness' and an elevated body temperature; and (iv) could not be stopped by voluntary effort." After extensive study of the subject, the experimenters concluded that "these data represent the clearest evidence to date that such a [CPG] network does exist in man."[66] Four years later, in 1998, Dimitrijevic, et al. showed that the human lumbar pattern generating networks can be activated by drive to large-diameter sensory afferents of the posterior roots.[10] When tonic electrical stimulation is applied to these fibers in motor complete spinal cord injured individuals (i.e., individuals in whom the spinal cord is functionally isolated from the brain) rhythmic, locomotor-like movement of the lower limbs can be elicited. These measurements were performed in supine position, thus minimizing peripheral feedback. Subsequent studies showed that these lumbar locomotor centers can form a large variety of rhythmic movements by combining and distributing stereotypical patterns to the numerous lower limb muscles.[11] A CPG-activating drug treatment called Spinalon, active centrally upon oral administration, has also been shown to partially reactivate spinal locomotor neurons in complete or motor-complete spinal cord-injured patients. Indeed, a double-blind, randomized, placebo-controlled study in forty-five volunteers with chronic AIS A/B injuries (between 3 months and 30 years post-trauma) lying in the supine position for safety reasons, revealed that Spinalon below maximum tolerated dose (MTD was 500/125/50 mg/kg L-DOPA/carbidopa/buspirone) was well tolerated. Preliminary evidence of efficacy was also found using videotape and electromyographic recordings since doses below MTD could acutely induce rhythmic locomotor-like leg movements in groups with Spinalon, but not in those with placebo (cornstarch).[67]
Neuromechanical control of locomotion in mammals
If step cycle durations and muscle activations were fixed, it wouldn't be possible to change body velocity and adapt to varying terrain. It has been suggested that the mammalian locomotor CPG comprises a “timer” (possibly in the form of coupled oscillators) which generates step cycles of varying durations, and a “pattern formation layer,” which selects and grades the activation of motor pools.[54][68] Increasing the neural drive from the midbrain locomotor region (MLR) to the spinal CPG increases the step cycle frequency (the cadence).[69] Swing and stance phase durations co-vary in a fairly fixed relationship, with stance phases changing more than swing phases.[70]
Sensory input from the limbs may truncate or extend individual phase durations in a process akin to finite state control (in which "if-then” rules determine when state transitions occur).[71][72][73] For example, if a limb that is swinging forward reaches the end of swing in less time than the current CPG-generated flexor phase duration, sensory input would cause the CPG timer to terminate swing and start the stance phase.[74][75] Furthermore, as body velocity increases, the pattern formation layer would increase muscle activation nonlinearly to provide increased load-bearing and thrust forces. It has been posited that in well-predicted movements, CPG-generated phase durations and muscle forces closely match those required by the evolving biomechanical events, minimizing the sensory corrections required. The term ‘‘neuromechanical tuning’’ has been coined to describe this process [57]
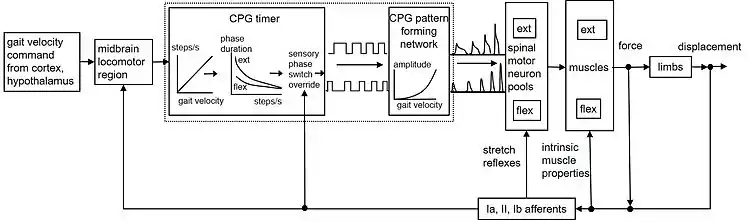
Fig. 1 provides a simplified schematic that summarizes these proposed mechanisms. A command specifying desired body velocity descends from higher centers to the MLR, which drives the spinal locomotor CPG. The CPG timer produces the appropriate cadence and phase durations and the pattern formation layer modulates the motoneuronal outputs.[76] The activated muscles resist stretch through their own intrinsic biomechanical properties, providing a rapid form of length and velocity feedback control. Reflexes mediated by Golgi tendon organ and other afferents provide additional load compensation, but the main role of sensory input may be to adjust or override the CPG at stance-swing-stance transitions.[77]
As described in Neuromodulation, the human locomotive CPG is very adaptable and can respond to sensory input. It receives input from the brainstem as well as from the environment to keep the network regulated. Newer studies have not only confirmed the presence of the CPG for human locomotion, but also confirmed its robustness and adaptability. For example, Choi and Bastian showed that the networks responsible for human walking are adaptable on short and long timescales. They showed adaptation to different gait patterns and different walking contexts. Also, they showed that different motor patterns can adapt independently. Adults could even walk on treadmills going in a different direction for each leg. This study showed that independent networks control forward and backward walking and that networks controlling each leg can adapt independently and be trained to walk independently.[78] Thus, humans also possess a central pattern generator for locomotion that is capable not only of rhythmic pattern generation but also remarkable adaptation and usefulness in a wide variety of situations.
Respiration
A three-phase model is the classical view of the respiratory CPG. The phases of the respiratory CPG are characterized by the rhythmic activity of: (1) the phrenic nerve during inspiration; (2) recurrent laryngeal nerve branches that innervate the thyroarytenoid muscle during the last stage of expiration; (3) the internal intercostal nerve branches that innervate the triangularis sterni muscle during the second stage of expiration. The rhythmicity of these nerves is classically viewed as originating from a single rhythm generator. In this model, phasing is produced by reciprocal synaptic inhibition between groups of sequentially active interneurons.
Nevertheless, an alternative model has been proposed[79] reinforced by certain experimental data. According to this model, respiratory rhythm is generated by two coupled anatomically distinct rhythm generators, one in the pre-Boetzinger complex[80] and the other in the retrotrapezoid nucleus / parafacial respiratory group. Further survey provided evidence to the hypothesis that one of the networks is responsible for inspiration rhythm and the other for expiration rhythm. Therefore, inspiration and expiration are distinct functions and one does not induce the other, as is the common belief, but one of two dominates the behavior by generating a faster rhythm.
Swallowing
Swallowing involves the coordinated contraction of more than 25 pairs of muscles in the oropharynx, larynx and esophagus, which are active during an oropharyngeal phase, followed by the primary esophageal peristalsis. Swallowing depends on a CPG located in the medulla oblongata, which involves several brain stem motor nuclei and two main groups of interneurons: a dorsal swallowing group (DSG) in the nucleus tractus solitarii and a ventral swallowing group (VSG) located in the ventrolateral medulla above the nucleus ambiguus. Neurons in the DSG are responsible for the generation of the swallowing pattern, while those in the VSG distribute the commands to the various motoneuronal pools. As in other CPGs, the functioning of the central network can be modulated by peripheral and central inputs, so that the swallowing pattern is adapted to the size of the bolus.
Within this network, central inhibitory connections play a major role, producing a rostrocaudal inhibition that parallels the rostrocaudal anatomy of the swallowing tract. Thus, when the neurons controlling the proximal parts of the tract are active, those that command more distal parts are inhibited. Apart from the type of connection between the neurons, intrinsic properties of the neurons, especially those of NTS neurons, probably also contribute to the shaping and timing of the swallowing pattern.
The swallowing CPG is a flexible CPG. This means that at least some of the swallowing neurons may be multifunctional neurons and belong to pools of neurons that are common to several CPGs. One such CPG is the respiratory one, which has been observed interacting with the swallowing CPG.[81][82]
Rhythm generators
Central pattern generators can also play a role in rhythm generation for other functions in vertebrates. For example, the rat vibrissa system uses an unconventional CPG for whisking movements. "Like other CPGs, the whisking generator can operate without cortical input or sensory feedback. However, unlike other CPGs, vibrissa motoneurons actively participate in rhythmogenesis by converting tonic serotonergic inputs into the patterned motor output responsible for movement of the vibrissae."[83] Breathing is another non-locomotive function of central pattern generators. For example, larval amphibians accomplish gas exchange largely through rhythmic ventilation of the gills. A study showed that lung ventilation in the tadpole brainstem may be driven by a pacemaker-like mechanism, whereas the respiratory CPG adapts in the adult bullfrog as it matures.[84] Thus, CPGs hold a broad range of functions in the vertebrate animal and are widely adaptable and variable with age, environment and behavior.
Mechanism of rhythmic generators: post-inhibitory rebound
Rhythmicity in CPG's can also result from time-dependent cellular properties such as adaptation, delayed excitation, and post-inhibitory rebound (PIR). PIR is an intrinsic property that elicits rhythmic electrical activity by depolarizing the membrane once hyperpolarizing stimulus is gone. It can be produced by several mechanisms including hyperpolarization-activated cation current (Ih), low-voltage activated calcium current,[85] or deinactivation of depolarization-activated inward currents.[86] Once inhibition has ceased, this period of PIR can be explained as the time with increased neuronal excitability. It is the property of many CNS neurons that sometimes results in action potential "bursts" following immediately after inhibitory synaptic input."Because of this, it has been suggested that PIR may contribute to the maintenance of oscillatory activity in neural networks that are characterized by mutual inhibitory connections, like those involved in locomotor behaviors. In addition, PIR is often included as an element in computational models of neural networks that involve mutual inhibition" [87] For example, the "PIR in crayfish stretch receptor neurons is caused by recovery from adaptation during the course of inhibitory hyperpolarization. One feature of that system is that PIR only occurs if the hyperpolarization is imposed on a background of excitation, caused in this case by stretch. They also found that PIR can be elicited in the stretch receptor by hyperpolarizing current pulses. This was an important finding because it showed that PIR is an intrinsic property of the postsynaptic neuron, related to the membrane potential change associated with inhibition but independent of transmitter receptors or presynaptic properties. The latter conclusion has stood the test of time, marking PIR as a robust property of CNS neurons in a wide variety of contexts."[88] This cellular property can most easily be seen in the Lamprey neural circuit. The swimming movement is produced by alternating neural activity between the left and right side of the body, causing it to bend back and forth while creating oscillating movements. While the Lamprey is bent to the left, there is reciprocal inhibition on the right side causing it to relax due to hyperpolarization. Immediately after this hyperopolarizing stimulus, the interneurons use post-inhibitory rebound to initiate activity in the right side. Depolarization of the membrane causes it to contract while reciprocal inhibition is now applied to the left side.
Functions in invertebrates
CPGs play a similarly critical role in coordinating behaviors in invertebrates, and studying invertebrate CPGs with fewer numbers of neurons has helped establish general principles of CPGs and their organization in the nervous system. One model circuit for studying CPGs is the stomatogastric ganglion in crabs and lobsters, a ~30 neuron circuit containing two CPGs that generate rhythmic motor output for chewing and digesting food.[7] Dissection of these circuits has revealed neural mechanisms of CPGs. For example, the pyloric CPG - which controls the contraction and dilation of the pylorus - contains a set of conditional oscillatory neurons and one pacemaker neuron that fires rhythmically when dissected out of the circuit.[7] Coordinated rhythmic behaviors like walking, flight and grooming are also controlled by CPGs in some invertebrates.[89][90][91][92] Continued research into how CPGs control these behaviors has revealed a nested CPG architecture to control rhythmic behaviors across various timescales.[93] Other examples of CPGs in invertebrate animals include a CPG modulating reflexive withdrawal, escape swimming and crawling in the mollusc Tritonia,[94] and to control the heartbeat of leeches.[95] Central pattern generators play a broad role in all animals and show amazing variability and adaptability in almost all cases.
Alternate Interpretations
One theory that reconciles the role of sensory feedback during rhythmic locomotion is to redefine CPGs as “state estimators” as opposed to rhythm generators[96] Through this perspective, CPGs are an intrinsic spinal processor that corrects imperfect sensory feedback and adapts central input to this optimized peripheral input.[97] Models using this framework are able to accomplish rhythmics behavior as well as fictive locomotion without incorporating independent rhythm generators.
References
- Wojcik, Jeremy; Clewley, Robert; Shilnikov, Andrey (16 May 2011). "Order parameter for bursting polyrhythms in multifunctional central pattern generators". Physical Review E. 83 (5): 056209. Bibcode:2011PhRvE..83e6209W. doi:10.1103/PhysRevE.83.056209. PMID 21728632.
- https://scholarworks.gsu.edu/cgi/viewcontent.cgi?article=1067&context=neurosci_facpub
- https://mathematical-neuroscience.springeropen.com/track/pdf/10.1186/s13408-018-0065-9.pdf
- Hooper, Scott L. (1999–2010). "Central Pattern Generators". Encyclopedia of Life Sciences. John Wiley & Sons. doi:10.1038/npg.els.0000032. ISBN 978-0-470-01590-2.
- Kuo 2002
- Guertin, PA. (January 2019). "Central pattern generators in the brainstem and spinal cord: an overview of basic principles, similarities and differences". Reviews in the Neurosciences. 30 (2): 107–164. doi:10.1515/revneuro-2017-0102. PMID 30543520. S2CID 56493287.
- Selverston, Allen I. (2010-08-12). "Invertebrate central pattern generator circuits". Philosophical Transactions of the Royal Society B: Biological Sciences. 365 (1551): 2329–2345. doi:10.1098/rstb.2009.0270. ISSN 0962-8436. PMC 2894947. PMID 20603355.
- Hultborn H, Nielsen JB (February 2007). "Spinal control of locomotion--from cat to man". Acta Physiologica. 189 (2): 111–21. doi:10.1111/j.1748-1716.2006.01651.x. PMID 17250563. S2CID 41080512.
- Guertin PA (December 2009). "The mammalian central pattern generator for locomotion". Brain Research Reviews. 62 (4): 345–56. doi:10.1016/j.brainresrev.2009.08.002. PMID 19720083. S2CID 9374670.
{{cite journal}}: CS1 maint: uses authors parameter (link) - Dimitrijevic MR, Gerasimenko Y, Pinter MM (November 1998). "Evidence for a spinal central pattern generator in humans". Annals of the New York Academy of Sciences. 860 (1): 360–76. Bibcode:1998NYASA.860..360D. doi:10.1111/j.1749-6632.1998.tb09062.x. PMID 9928325. S2CID 102514.
- Danner SM, Hofstoetter US, Freundl B, Binder H, Mayr W, Rattay F, Minassian K (March 2015). "Human spinal locomotor control is based on flexibly organized burst generators". Brain. 138 (Pt 3): 577–88. doi:10.1093/brain/awu372. PMC 4408427. PMID 25582580.
- Minassian, Karen; Hofstoetter, Ursula S.; Dzeladini, Florin; Guertin, Pierre A.; Ijspeert, Auke (2017). "The Human Central Pattern Generator for Locomotion: Does It Exist and Contribute to Walking?". The Neuroscientist. 23 (6): 649–663. doi:10.1177/1073858417699790. PMID 28351197. S2CID 33273662.
- Marder, Eve; Bucher, Dirk (2001-11-27). "Central pattern generators and the control of rhythmic movements". Current Biology. 11 (23): R986–R996. doi:10.1016/S0960-9822(01)00581-4. ISSN 0960-9822. PMID 11728329. S2CID 1294374.
- Marder, E.; Eisen, J. S. (June 1984). "Electrically coupled pacemaker neurons respond differently to same physiological inputs and neurotransmitters". Journal of Neurophysiology. 51 (6): 1362–1374. doi:10.1152/jn.1984.51.6.1362. ISSN 0022-3077. PMID 6145758.
- Smith, J. C.; Ellenberger, H. H.; Ballanyi, K.; Richter, D. W.; Feldman, J. L. (1991-11-01). "Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals". Science. 254 (5032): 726–729. Bibcode:1991Sci...254..726S. doi:10.1126/science.1683005. ISSN 0036-8075. PMC 3209964. PMID 1683005.
- Wang, Xiao-Jing; Rinzel, John (January 1992). "Alternating and Synchronous Rhythms in Reciprocally Inhibitory Model Neurons". Neural Computation. 4 (1): 84–97. doi:10.1162/neco.1992.4.1.84. ISSN 0899-7667. S2CID 33448885.
- Brown, T. Graham (1914-03-31). "On the nature of the fundamental activity of the nervous centres; together with an analysis of the conditioning of rhythmic activity in progression, and a theory of the evolution of function in the nervous system". The Journal of Physiology. 48 (1): 18–46. doi:10.1113/jphysiol.1914.sp001646. ISSN 0022-3751. PMC 1420503. PMID 16993247.
- Satterlie, Richard A. (1985-07-26). "Reciprocal Inhibition and Postinhibitory Rebound Produce Reverberation in a Locomotor Pattern Generator". Science. 229 (4711): 402–404. Bibcode:1985Sci...229..402S. doi:10.1126/science.229.4711.402. PMID 17795901. S2CID 44315274.
- Roberts, A.; Soffe, S. R.; Wolf, E. S.; Yoshida, M.; Zhao, F. Y. (1998-11-16). "Central circuits controlling locomotion in young frog tadpoles". Annals of the New York Academy of Sciences. 860 (1): 19–34. Bibcode:1998NYASA.860...19R. doi:10.1111/j.1749-6632.1998.tb09036.x. ISSN 0077-8923. PMID 9928299. S2CID 24563208.
- Grillner, S.; Wallén, P. (1999). On the cellular bases of vertebrate locomotion. Progress in Brain Research. Vol. 123. pp. 297–309. doi:10.1016/s0079-6123(08)62865-4. ISSN 0079-6123. PMID 10635725.
- Nadim, Farzan; Manor, Yair (2000-12-01). "The role of short-term synaptic dynamics in motor control". Current Opinion in Neurobiology. 10 (6): 683–690. doi:10.1016/S0959-4388(00)00159-8. ISSN 0959-4388. PMID 11240276. S2CID 19397634.
- Bucher, Dirk; Haspel, Gal; Golowasch, Jorge; Nadim, Farzan (2015-12-23). "Central Pattern Generators". eLS: 1–12. doi:10.1002/9780470015902.a0000032.pub2. ISBN 9780470016176.
- Li, Wen-Chang; Sautois, Bart; Roberts, Alan; Soffe, Stephen R. (2007-11-07). "Reconfiguration of a Vertebrate Motor Network: Specific Neuron Recruitment and Context-Dependent Synaptic Plasticity". Journal of Neuroscience. 27 (45): 12267–12276. doi:10.1523/JNEUROSCI.3694-07.2007. ISSN 0270-6474. PMC 6673254. PMID 17989292.
- Rubin, Jonathan E.; Hayes, John A.; Mendenhall, Jeffrey L.; Del Negro, Christopher A. (2009-02-24). "Calcium-activated nonspecific cation current and synaptic depression promote network-dependent burst oscillations". Proceedings of the National Academy of Sciences of the United States of America. 106 (8): 2939–2944. Bibcode:2009PNAS..106.2939R. doi:10.1073/pnas.0808776106. ISSN 0027-8424. PMC 2636730. PMID 19196976.
- Kiehn O, Butt SJ (July 2003). "Physiological, anatomical and genetic identification of CPG neurons in the developing mammalian spinal cord". Prog. Neurobiol. 70 (4): 347–61. doi:10.1016/S0301-0082(03)00091-1. PMID 12963092. S2CID 22793900.
- Bidaye, Salil S.; Bockemühl, Till; Büschges, Ansgar (2018-02-01). "Six-legged walking in insects: how CPGs, peripheral feedback, and descending signals generate coordinated and adaptive motor rhythms". Journal of Neurophysiology. 119 (2): 459–475. doi:10.1152/jn.00658.2017. ISSN 1522-1598. PMID 29070634.
- Jean A (April 2001). "Brain stem control of swallowing: neuronal network and cellular mechanisms". Physiol. Rev. 81 (2): 929–69. doi:10.1152/physrev.2001.81.2.929. PMID 11274347. S2CID 17609854.
- Cunningham ET, Sawchenko PE (February 2000). "Dorsal medullary pathways subserving oromotor reflexes in the rat: implications for the central neural control of swallowing". J. Comp. Neurol. 417 (4): 448–66. doi:10.1002/(SICI)1096-9861(20000221)417:4<448::AID-CNE5>3.0.CO;2-S. PMID 10701866. S2CID 2403930.
- Goulding, Martyn (July 2009). "Circuits controlling vertebrate locomotion: moving in a new direction". Nature Reviews Neuroscience. 10 (7): 507–518. doi:10.1038/nrn2608. ISSN 1471-0048. PMC 2847453. PMID 19543221.
- Berg, Eva M.; Björnfors, E. Rebecka; Pallucchi, Irene; Picton, Laurence D.; El Manira, Abdeljabbar (2018). "Principles Governing Locomotion in Vertebrates: Lessons From Zebrafish". Frontiers in Neural Circuits. 12: 73. doi:10.3389/fncir.2018.00073. ISSN 1662-5110. PMC 6146226. PMID 30271327.
- Jessell, Thomas M. (October 2000). "Neuronal specification in the spinal cord: inductive signals and transcriptional codes". Nature Reviews Genetics. 1 (1): 20–29. doi:10.1038/35049541. ISSN 1471-0064. PMID 11262869. S2CID 205012382.
- Gosgnach, Simon; Bikoff, Jay B.; Dougherty, Kimberly J.; El Manira, Abdeljabbar; Lanuza, Guillermo M.; Zhang, Ying (2017-11-08). "Delineating the Diversity of Spinal Interneurons in Locomotor Circuits". The Journal of Neuroscience. 37 (45): 10835–10841. doi:10.1523/JNEUROSCI.1829-17.2017. ISSN 1529-2401. PMC 6596484. PMID 29118212.
- Rybak, Ilya A.; Dougherty, Kimberly J.; Shevtsova, Natalia A. (2015-09-22). "Organization of the Mammalian Locomotor CPG: Review of Computational Model and Circuit Architectures Based on Genetically Identified Spinal Interneurons". eNeuro. 2 (5): ENEURO.0069–15.2015. doi:10.1523/ENEURO.0069-15.2015. ISSN 2373-2822. PMC 4603253. PMID 26478909.
- Harris-Warrick, Ronald M. (October 2011). "Neuromodulation and Flexibility in Central Pattern Generator Networks". Current Opinion in Neurobiology. 21 (5): 685–692. doi:10.1016/j.conb.2011.05.011. ISSN 0959-4388. PMC 3171584. PMID 21646013.
- Katz, Paul S (1995-12-01). "Intrinsic and extrinsic neuromodulation of motor circuits". Current Opinion in Neurobiology. 5 (6): 799–808. doi:10.1016/0959-4388(95)80109-X. ISSN 0959-4388. PMID 8805409. S2CID 43818751.
- Schmidt, Brian J; Jordan, Larry M (2000-11-15). "The role of serotonin in reflex modulation and locomotor rhythm production in the mammalian spinal cord". Brain Research Bulletin. 53 (5): 689–710. doi:10.1016/S0361-9230(00)00402-0. ISSN 0361-9230. PMID 11165804. S2CID 20952920.
- Nadim, Farzan; Bucher, Dirk (2014-12-01). "Neuromodulation of neurons and synapses". Current Opinion in Neurobiology. SI: Neuromodulation. 29: 48–56. doi:10.1016/j.conb.2014.05.003. ISSN 0959-4388. PMC 4252488. PMID 24907657.
- Marder, Eve (2012-10-04). "Neuromodulation of Neuronal Circuits: Back to the Future". Neuron. 76 (1): 1–11. doi:10.1016/j.neuron.2012.09.010. ISSN 0896-6273. PMC 3482119. PMID 23040802.
- Blitz, Dawn M.; Christie, Andrew E.; Coleman, Melissa J.; Norris, Brian J.; Marder, Eve; Nusbaum, Michael P. (1999-07-01). "Different Proctolin Neurons Elicit Distinct Motor Patterns from a Multifunctional Neuronal Network". Journal of Neuroscience. 19 (13): 5449–5463. doi:10.1523/JNEUROSCI.19-13-05449.1999. ISSN 0270-6474. PMC 6782314. PMID 10377354.
- Harris-Warrick RM, Johnson BR, Peck JH, Kloppenburg P, Ayali A, Skarbinski J (November 1998). "Distributed effects of dopamine modulation in the crustacean pyloric network". Annals of the New York Academy of Sciences. 860 (1 NeuronaL Mech): 155–67. Bibcode:1998NYASA.860..155H. doi:10.1111/j.1749-6632.1998.tb09046.x. PMID 9928309. S2CID 23623832.
- Harris-Warrick, Ronald M. (October 2011). "Neuromodulation and Flexibility in Central Pattern Generator Networks". Current Opinion in Neurobiology. 21 (5): 685–692. doi:10.1016/j.conb.2011.05.011. ISSN 0959-4388. PMC 3171584. PMID 21646013.
- Dickinson, Patsy S.; Mecsas, Carol; Marder, Eve (March 1990). "Neuropeptide fusion of two motor-pattern generator circuits". Nature. 344 (6262): 155–158. Bibcode:1990Natur.344..155D. doi:10.1038/344155a0. ISSN 1476-4687. PMID 2308633. S2CID 4260725.
- Hooper, Scott L.; Moulins, Maurice (1989-06-30). "Switching of a Neuron from One Network to Another by Sensory-Induced Changes in Membrane Properties". Science. 244 (4912): 1587–1589. Bibcode:1989Sci...244.1587H. doi:10.1126/science.2740903. PMID 2740903.
- Gottschall JS, Nichols TR (September 2007). "Head pitch affects muscle activity in the decerebrate cat hindlimb during walking". Exp Brain Res. 182 (1): 131–5. doi:10.1007/s00221-007-1084-z. PMC 3064865. PMID 17690872.
- Graham-Brown, T. (1911). "The intrinsic factors in the act of progression in the mammal". Philosophical Transactions of the Royal Society of London B. 84 (572): 308–319. Bibcode:1911RSPSB..84..308B. doi:10.1098/rspb.1911.0077.
- Whelan PJ (December 2003). "Developmental aspects of spinal locomotor function: insights from using the in vitro mouse spinal cord preparation". J. Physiol. 553 (Pt 3): 695–706. doi:10.1113/jphysiol.2003.046219. PMC 2343637. PMID 14528025.
- Ayers J, Carpenter GA, Currie S, Kinch J (September 1983). "Which behavior does the lamprey central motor program mediate?". Science. 221 (4617): 1312–4. Bibcode:1983Sci...221.1312A. doi:10.1126/science.6137060. PMID 6137060.
- Harris-Warrick R, Cohen A (1985) Serotonin modulates the central pattern generator for locomotion in the isolated lamprey spinal cord. J Exp Biol 116:27-46.
- Parker D, Zhang W, Grillner S (1998). "Substance P modulates NMDA responses and causes long-term protein synthesis-dependent modulation of the lamprey locomotor network". J Neurosci. 18 (12): 4800–4813. doi:10.1523/JNEUROSCI.18-12-04800.1998. PMC 6792700. PMID 9614253.
- Parker D (January 2006). "Complexities and uncertainties of neuronal network function". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 361 (1465): 81–99. doi:10.1098/rstb.2005.1779. PMC 1626546. PMID 16553310.
- Parker D (August 2010). "Neuronal network analyses: premises, promises and uncertainties". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 365 (1551): 2315–28. doi:10.1098/rstb.2010.0043. PMC 2894952. PMID 20603354.
- Zhaoping, Li; Lewis, Alex; Scarpetta, Silvia (2004-05-14). "Mathematical Analysis and Simulations of the Neural Circuit for Locomotion in Lampreys". Physical Review Letters. 92 (19): 198106. arXiv:q-bio/0404012. Bibcode:2004PhRvL..92s8106Z. doi:10.1103/PhysRevLett.92.198106. PMID 15169452. S2CID 7790379.
- Ijspeert, Auke Jan and Jerome Kodjabachian "Evolution and development of a central pattern generator for the swimming of a lamprey." Research Paper No 926, Dept. of Artificial Intelligence, University of Edinburgh, 1998
- Rybak IA, Shevtsova NA, Lafreniere-Roula M, McCrea DA (December 2006). "Modelling spinal circuitry involved in locomotor pattern generation: insights from deletions during fictive locomotion". The Journal of Physiology. 577 (Pt 2): 617–39. doi:10.1113/jphysiol.2006.118703. PMC 1890439. PMID 17008376.
- Bashor DP, Dai Y, Kriellaars DJ, Jordan LM (November 1998). "Pattern generators for muscles crossing more than one joint". Annals of the New York Academy of Sciences. 860 (1 Neuronal Mech): 444–7. Bibcode:1998NYASA.860..444B. CiteSeerX 10.1.1.215.3329. doi:10.1111/j.1749-6632.1998.tb09071.x. PMID 9928334. S2CID 7322093.
- Yakovenko S, McCrea DA, Stecina K, Prochazka A (August 2005). "Control of locomotor cycle durations". Journal of Neurophysiology. 94 (2): 1057–65. CiteSeerX 10.1.1.215.8127. doi:10.1152/jn.00991.2004. PMID 15800075.
- Prochazka A, Yakovenko S (2007). "The neuromechanical tuning hypothesis". Computational Neuroscience: Theoretical Insights into Brain Function. Prog. Brain Res. Progress in Brain Research. Vol. 165. pp. 255–65. doi:10.1016/S0079-6123(06)65016-4. ISBN 9780444528230. PMID 17925251.
- Danner SM, Wilshin SD, Shevtsova NA, Rybak IA (December 2016). "Central control of interlimb coordination and speed-dependent gait expression in quadrupeds". The Journal of Physiology. 594 (23): 6947–6967. doi:10.1113/JP272787. PMC 5134391. PMID 27633893.
- Talpalar AE, Bouvier J, Borgius L, Fortin G, Pierani A, Kiehn O (August 2013). "Dual-mode operation of neuronal networks involved in left-right alternation". Nature. 500 (7460): 85–8. Bibcode:2013Natur.500...85T. doi:10.1038/nature12286. PMID 23812590. S2CID 4427401.
- Kiehn O (April 2016). "Decoding the organization of spinal circuits that control locomotion". Nature Reviews. Neuroscience. 17 (4): 224–38. doi:10.1038/nrn.2016.9. PMC 4844028. PMID 26935168.
- Danner SM, Shevtsova NA, Frigon A, Rybak IA (November 2017). "Computational modeling of spinal circuits controlling limb coordination and gaits in quadrupeds". eLife. 6. doi:10.7554/eLife.31050. PMC 5726855. PMID 29165245.
- Ausborn, Jessica; Shevtsova, Natalia A.; Danner, Simon M. (25 June 2021). "Computational Modeling of Spinal Locomotor Circuitry in the Age of Molecular Genetics". International Journal of Molecular Sciences. 22 (13): 6835. doi:10.3390/ijms22136835. PMC 8267724. PMID 34202085.
- Bellardita C, Kiehn O (June 2015). "Phenotypic characterization of speed-associated gait changes in mice reveals modular organization of locomotor networks". Current Biology. 25 (11): 1426–36. doi:10.1016/j.cub.2015.04.005. PMC 4469368. PMID 25959968.
- Ruder L, Takeoka A, Arber S (December 2016). "Long-Distance Descending Spinal Neurons Ensure Quadrupedal Locomotor Stability". Neuron. 92 (5): 1063–1078. doi:10.1016/j.neuron.2016.10.032. PMID 27866798.
- Ausborn J, Shevtsova NA, Caggiano V, Danner SM, Rybak IA (January 2019). "Computational modeling of brainstem circuits controlling locomotor frequency and gait". eLife. 8. doi:10.7554/eLife.43587. PMC 6355193. PMID 30663578.
- Calancie B, Needham-Shropshire B, Jacobs P, Willer K, Zych G, Green BA (October 1994). "Involuntary stepping after chronic spinal cord injury. Evidence for a central rhythm generator for locomotion in man". Brain. 117 (Pt 5): 1143–59. doi:10.1093/brain/117.5.1143. PMID 7953595.
- Radhakrishna M, Steuer I, Prince F, Roberts M, Mongeon D, Kia M, Dyck S, Matte G, Vaillancourt M, Guertin PA (December 2017). "Double-blind, placebo-controlled, randomized phase I/IIa study (safety and efficacy) with buspirone/levodopa/carbidopa (Spinalon) in subjects with complete AIS A or motor-complete AIS B spinal cord injury". Current Pharmaceutical Design. 23 (12): 1789–1804. doi:10.2174/1381612822666161227152200. PMID 28025945.
- Perret C, Cabelguen JM (1980). "Main characteristics of the hindlimb locomotor cycle in the decorticate cat with special reference to bifunctional muscles". Brain Research. 187 (2): 333–352. doi:10.1016/0006-8993(80)90207-3. PMID 7370734. S2CID 44913308.
- Shik ML, Severin FV, Orlovsky GN (1966). "Control of walking and running by means of electrical stimulation of the mid-brain". Biophysics. 11: 756–765.
- Goslow GE Jr.; Reinking RM; Stuart DG (1973). "The cat step cycle: hind limb joint angles and muscle lengths during unrestrained locomotion". Journal of Morphology. 141 (1): 1–41. doi:10.1002/jmor.1051410102. PMID 4727469. S2CID 42918929.
- Cruse H (1990). "What mechanisms coordinate leg movement in walking arthropods?" (PDF). Trends in Neurosciences. 13 (1): 15–21. doi:10.1016/0166-2236(90)90057-h. PMID 1688670. S2CID 16401306.
- Hemami H, Tomovic R, Ceranowicz AZ (1978). "Finite state control of planar bipeds with application to walking and sitting". Journal of Bioengineering. 2 (6): 477–494. PMID 753838.
- Prochazka A (1993). "Comparison of natural and artificial control of movement". IEEE Trans Rehab Eng. 1: 7–17. doi:10.1109/86.242403.
- Hiebert GW, Whelan PJ, Prochazka A, Pearson KG (1996). "Contribution of hind limb flexor muscle afferents to the timing of phase transitions in the cat step cycle". Journal of Neurophysiology. 75 (3): 1126–1137. doi:10.1152/jn.1996.75.3.1126. PMID 8867123.
- Guertin P, Angel MJ, Perreault MC, McCrea DA (1995). "Ankle extensor group I afferents excite extensors throughout the hindlimb during fictive locomotion in the cat". Journal of Physiology. 487 (1): 197–209. doi:10.1113/jphysiol.1995.sp020871. PMC 1156609. PMID 7473249.
- Prochazka A, Ellaway PH (2012). "Sensory systems in the control of movement". Comprehensive Physiology, Supplement 29: Handbook of Physiology, Exercise: Regulation and Integration of Multiple Systems. NY: John Wiley & Sons in conjunction with the American Physiological Society. pp. 2615‐2627.
- Donelan JM, McVea DA, Pearson KG (2009). "Force regulation of ankle extensor muscle activity in freely walking cats". J Neurophysiol. 101 (1): 360–371. doi:10.1152/jn.90918.2008. PMID 19019974.
- Choi JT, Bastian AJ (August 2007). "Adaptation reveals independent control networks for human walking". Nat. Neurosci. 10 (8): 1055–62. doi:10.1038/nn1930. PMID 17603479. S2CID 1514215.
- Janczewski WA, Feldman JL (January 2006). "Distinct rhythm generators for inspiration and expiration in the juvenile rat". The Journal of Physiology. 570 (Pt 2): 407–20. doi:10.1113/jphysiol.2005.098848. PMC 1464316. PMID 16293645.
- Smith JC, Ellenberger HH, Ballanyi K, Richter DW, Feldman JL (November 1991). "Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals". Science. 254 (5032): 726–9. Bibcode:1991Sci...254..726S. doi:10.1126/science.1683005. PMC 3209964. PMID 1683005.
- Dick TE, Oku Y, Romaniuk JR, Cherniack NS (June 1993). "Interaction between central pattern generators for breathing and swallowing in the cat". The Journal of Physiology. 465: 715–30. doi:10.1113/jphysiol.1993.sp019702. PMC 1175455. PMID 8229859.
- Grélot L, Barillot JC, Bianchi AL (1989). "Pharyngeal motoneurones: respiratory-related activity and responses to laryngeal afferents in the decerebrate cat". Experimental Brain Research. 78 (2): 336–44. doi:10.1007/bf00228905. PMID 2599043. S2CID 605299.
- Cramer NP, Li Y, Keller A (March 2007). "The whisking rhythm generator: a novel mammalian network for the generation of movement". Journal of Neurophysiology. 97 (3): 2148–58. doi:10.1152/jn.01187.2006. PMC 1821005. PMID 17202239.
- Broch L, Morales RD, Sandoval AV, Hedrick MS (April 2002). "Regulation of the respiratory central pattern generator by chloride-dependent inhibition during development in the bullfrog (Rana catesbeiana)". The Journal of Experimental Biology. 205 (Pt 8): 1161–9. doi:10.1242/jeb.205.8.1161. PMID 11919275.
- Fan, Yi-Ping; Horn, Eric M.; Waldrop, Tony G. (2000-12-01). "Biophysical Characterization of Rat Caudal Hypothalamic Neurons: Calcium Channel Contribution to Excitability". Journal of Neurophysiology. 84 (6): 2896–2903. doi:10.1152/jn.2000.84.6.2896. ISSN 0022-3077. PMID 11110819. S2CID 1753434.
- Angstadt JD, Grassmann JL, Theriault KM, Levasseur SM (August 2005). "Mechanisms of postinhibitory rebound and its modulation by serotonin in excitatory swim motor neurons of the medicinal leech". Journal of Comparative Physiology A. 191 (8): 715–32. doi:10.1007/s00359-005-0628-6. PMID 15838650. S2CID 31433117.
- Perkel DH, Mulloney B (July 1974). "Motor pattern production in reciprocally inhibitory neurons exhibiting postinhibitory rebound". Science. 185 (4146): 181–3. Bibcode:1974Sci...185..181P. doi:10.1126/science.185.4146.181. PMID 4834220. S2CID 38173947.
- Gerasimov VD, Kostyuk PG, Maiskii VA (1966). "Reactions of giant neurons to break of hyperpolarizing current". Federation Proceedings. Translation Supplement; Selected Translations from Medical-related Science. 25 (3): 438–42. PMID 5222090.
- Mantziaris, Charalampos; Bockemühl, Till; Büschges, Ansgar (2020). "Central pattern generating networks in insect locomotion". Developmental Neurobiology. 80 (1–2): 16–30. doi:10.1002/dneu.22738. ISSN 1932-846X. PMID 32128970. S2CID 212407130.
- Rillich, Jan; Stevenson, Paul A.; Pflueger, Hans-Joachim (2013-05-09). "Flight and Walking in Locusts–Cholinergic Co-Activation, Temporal Coupling and Its Modulation by Biogenic Amines". PLOS ONE. 8 (5): e62899. Bibcode:2013PLoSO...862899R. doi:10.1371/journal.pone.0062899. ISSN 1932-6203. PMC 3650027. PMID 23671643.
- Berkowitz, A.; Laurent, G. (1996-12-15). "Central generation of grooming motor patterns and interlimb coordination in locusts". The Journal of Neuroscience. 16 (24): 8079–8091. doi:10.1523/JNEUROSCI.16-24-08079.1996. ISSN 0270-6474. PMC 6579234. PMID 8987833.
- Marder, E.; Bucher, D. (2001-11-27). "Central pattern generators and the control of rhythmic movements". Current Biology. 11 (23): R986–996. doi:10.1016/s0960-9822(01)00581-4. ISSN 0960-9822. PMID 11728329. S2CID 1294374.
- Ravbar, Primoz; Zhang, Neil; Simpson, Julie H. (2021-07-05). "Behavioral evidence for nested central pattern generator control of Drosophila grooming". eLife. 10: 2020.09.15.298679. bioRxiv 10.1101/2020.09.15.298679. doi:10.7554/eLife.71508. PMC 8694699. PMID 34936550. S2CID 221823475.
- Popescu IR, Frost WN (March 2002). "Highly dissimilar behaviors mediated by a multifunctional network in the marine mollusk Tritonia diomedea". J. Neurosci. 22 (5): 1985–93. doi:10.1523/JNEUROSCI.22-05-01985.2002. PMC 6758888. PMID 11880529.
- Norris, Brian J.; Weaver, Adam L.; Wenning, Angela; García, Paul S.; Calabrese, Ronald L. (2007-11-01). "A Central Pattern Generator Producing Alternative Outputs: Phase Relations of Leech Heart Motor Neurons With Respect to Premotor Synaptic Input". Journal of Neurophysiology. 98 (5): 2983–2991. doi:10.1152/jn.00407.2007. ISSN 0022-3077. PMID 17728387.
- Ryu, Hansol X., and Arthur D. Kuo. "An optimality principle for locomotor central pattern generators." bioRxiv (2019)
- Kuo 2002.
External links
- How Do Central Pattern Generators Work? @ www.bio.brandeis.edu
- Hooper's Review of Central Pattern Generators(PDF)
- Dimitrijevic MR, Gerasimenko Y, Pinter MM (November 1998). "Evidence for a spinal central pattern generator in humans". Annals of the New York Academy of Sciences. 860 (1 Neuronal Mech): 360–76. Bibcode:1998NYASA.860..360D. doi:10.1111/j.1749-6632.1998.tb09062.x. PMID 9928325. S2CID 102514.
- Ijspeert AJ, Kodjabachian J (1998). "Evolution and development of a central pattern generator for the swimming of a lamprey". Artificial Life. Dept. of Artificial Intelligence, University of Edinburgh. 5 (3): 247–69. doi:10.1162/106454699568773. PMID 10648954. S2CID 14863127. Research Paper No. 926.
- Kuo AD (April 2002). "The relative roles of feedforward and feedback in the control of rhythmic movements" (PDF). Motor Control. 6 (2): 129–45. doi:10.1123/mcj.6.2.129. PMID 12122223.
- Katz PS, Fickbohm DJ, Lynn-Bullock CP (2001). "Evidence that the Central Pattern Generator for Swimming in Tritonia Arose from a Non-Rhythmic Neuromodulatory Arousal System: Implications for the Evolution of Specialized Behavior". Am. Zool. 41 (4): 962–975. CiteSeerX 10.1.1.526.3368. doi:10.1093/icb/41.4.962.
- Carew J.T. (2000). Behavioral Neurobiology. Sunderland MA: Sinauer Associates. pp. 155–163.
| Concepts |
|
|---|---|
| People |
|
| Methods | |
| Systems |
|
| |